
I canali ionici sono proteine presenti nella membrana cellulare che permettono il passaggio selettivo di ioni attraverso la membrana. Questi canali sono importanti per la regolazione del potenziale di membrana e per la trasmissione degli impulsi nervosi. I canali ionici possono essere regolati da diversi fattori, come il voltaggio, i ligandi o gli stimoli meccanici. La loro attività è influenzata anche dai gradienti di concentrazione ionica. Esistono diversi tipi di canali ionici, come i canali del sodio, del potassio, del calcio e del cloro. La loro funzione è essenziale per il corretto funzionamento delle cellule e degli organismi.
Nel corso di un potenziale di azione, il potenziale di membrana può andare incontro a rapide variazioni, esse sono rese possibili dalla presenza di canali ionici che costituiscono una classe di proteine integrali che attraversano la membrana da parte a parte. Questi canali possiedono tre proprietà importanti:
- Lasciano passare gli ioni
- Riconoscono e selezionano specie ioniche
- .Si aprono e si chiudono in risposta a segnali specifici che possono essere di natura elettrica, chimica o meccanica.
A riposo, il potenziale di membrana delle cellule nervose è determinato in gran parte da canali ionici permeabili al potassio (K), ma nel corso di un potenziale di azione vengono attivati canali selettivi per il sodio (Na). Ogni canale ionico possiede almeno uno strato aperto e uno o due strati di chiusura. I canali nei quali si può avere questa transizione di strato vengono detti canali ad accesso variabile. Le variazioni d’accesso di un canale comportano modificazioni diffuse di tutta la sua struttura, si pensa che l’apertura e la chiusura di questi canali comporti l’inclinazione e la rotazione contemporanea delle sei subunità che costituiscono il canale.
I canali passivi che sono generalmente sempre aperti, rivestono un importanza notevole nel mantenimento del potenziale a riposo in quanto consentono il passaggio degli ioni mantenendo il gradiente di concentrazione.
Alternativamente i canali possono essere regolati da tre meccanismi diversi:
- dal voltaggio (canali voltaggio dipendenti), come quello per il sodio (Na), l’apertura e la chiusura del canale è legata dalle variazioni del voltaggio esistente ai capi della membrana e ciò determina la transizione del canale tra gli stati chiuso e aperto.
- da trasmettitori chimici (canali dipendenti da ligandi), i ligandi possono essere neurotrasmettitori o ormoni che si trovano negli spazi extracellulari, o da secondi messaggeri intracellulari attivati da neurotrasmettitori.
- e da pressione o stiramento (canali dipendenti da stimoli meccanici) si ritiene che sia il citoscheletro che trasferisce sul canale l’energia associata allo stiramento.
Sotto l’influenza di questi fattori i canali ionici possono presentarsi in tre stati funzionali diversi: chiuso ma attivabile (di riposo), aperto (attivo), chiuso e non attivabile (refrattario).
I canali quando si aprono tendono a rimanere aperti per alcuni millesecondi, e analogamente, quando si chiudono, rimangono chiusi alcuni millesecondi prima di riaprirsi, questa transizione da inizio ad una brusca variazione a gradino e con carattere di tutto o nulla.
I canali dipendenti da ligandi possono entrare in stato di refrattarietà quando vengono esposti a lungo all’azione del ligando stesso. Questo processo viene detto di desensibilizzazione (o adattamento) e in alcuni casi sembra essere una proprietà intrinseca del canale, mentre in altri casi è dovuta alla fosforilazione del canale dovuta ad una protein-chinasi.
Per ciò che riguarda i canali voltaggio dipendenti, molti ma non tutti, entrano in una fase di refrattarietà dopo essere stati attivati. Questo processo è detto stato di inattivazione. Si ritiene che l’inattivazione dei canali Na e K voltaggio dipendente sia dovuta ad un cambio di conformazione sotto il controllo di una subunità diversa da quella che determina la loro attivazione. Anche i fattori esogeni come le tossine possono influenzare i siti che controllano l’accesso dei canali ionici. Alcune sostanze si legano allo stesso sito al quale normalmente si associa il ligando che determina l’apertura del canale, impedendo in tal modo all’attivatore di esercitare il proprio effetto.
I canali ionici sono proteine di grandi dimensioni, coniugate in superficie con carboidrati (glicoproteine) che attraversano parte a parte le membrane. Tutti i canali hanno un poro idrofilo centrale che attraversa la membrana a tutto spessore.
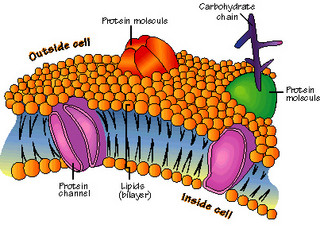
La membrana è costituita da uno doppio strato fosfolipidico. I lipidi delle membrane sono idrofobici e non sono miscibili con l’acqua. L’atomo di ossigeno della molecola di acqua tende ad attrarre elettroni e porta perciò una piccola carica negativa netta, mentre gli atomi di idrogeno (H) tendono a perdere elettroni ed acquistano quindi una piccola carica positiva netta. Per via di questa distribuzione di cariche, i cationi subiscono una attrazione da parte dell’atomo di ossigeno (O) mentre gli anioni sono attratti da quelli di H. Per questo motivo gli ioni dei lipidi idrofili attraggono l’acqua, e ciò provoca una nuvola di molecole di acqua di idratazione legata elettrostaticamente.
Affinchè uno ione possa uscire dalla fase acquosa ed entrare nel doppio strato lipidico delle membrane è necessario che gli sia fornita una gran quantità di energia per vincere le forze di attrazione che si stabiliscono tra gli ioni e le molecole di acqua circostanti.
Gli ioni possono dunque attraversare la membrana attraverso pori o aperture specializzate della membrana, come i canali ionici all’interno dei quali il bilancio energetico è orientato in modo da favorire il flusso degli ioni.
Da queste premesse ci si può porre il quesito di come gli ioni si spostino attraverso i canali. In una soluzione di acqua di idratazione la facilità con cui uno ione si muove ( la costante di diffusione) è in relazione alle sue dimensioni ioniche, tanto può piccolo è uno ione tanto maggiormente localizzata è la sua carica.
Di conseguenza uno ione piccolo come il Na possiede un campo elettrico maggiore di uno ione di dimensioni maggiori come quello del K, perché gli ioni piccoli esercitano un’attrazione maggiore sulle molecole di acqua. Così quando uno ione Na si muove nella soluzione acquosa la maggiore attrazione che esso esercita sulle molecole di acqua lo faranno rallentare rispetto ad un K. Ciò dimostra come un canale possa essere selettivo per il K. Il problema opposto è invece come un canale può essere selettivo per il Na . Oggi sappiamo dell’esistenza della pompa Na – K.

Per spiegare la selettività la teoria poro-canali ipotizza che all’interno del canale vi sia un filtro di selettività, nel quale gli ioni cederebbero la maggior parte della loro acqua di idratazione. Questa perdita rappresenta una condizione sfavorevole per lo ione, quindi esso riuscirà a passare se l’energia acquistata mediante l’interazione con il filtro di selettività riuscirà a compensare quella perduta con l’acqua d’idratazione.
L’intensità della corrente di un singolo canale può venire misurata con la legge di Ohm, i=V/R dove i è la corrente che passa attraverso il canale, V il voltaggio e R la resistenza.

Le proprietà elettriche dei canali possono essere confermate con la tecnica del patch-clamp. Se si porta in stretta opposizione con la membrana di una fibra muscolare di rana una micropipetta di vetro riempita di acetilcolina (Ach), nella zona sottostante la punta della micropipetta è possibile registrare impulsi unitari di corrente (potenziali di placca in miniatura), che rappresentano la chiusura e l’apertura dei singoli canali ionici.
Le cinetiche dei flussi ionici attraverso i canali sono caratterizzati dalla conduttanza in risposta ad una certa forza elettrochimica. La forza elettrochimica netta è determinata da due fattori: dalla differenza di potenziale e dal gradiente di concentrazione dello ione considerato presenti ai capi della membrana.
In alcuni canali il flusso di corrente varia linearmente con la forza elettromotrice ed i canali si comportano come delle semplici resistenze (vedi esempio di un circuito equivalente del neurone). In altri canali, il flusso di corrente non è lineare con la forza elettromotrice. Questi canali si comportano come dei rettificatori di corrente, cioè come delle componenti che trasportano maggiormente gli ioni in una direzione che non in quella contraria.
I flussi ionici che attraversano i canali dipendono dalle concentrazioni ioniche presenti nelle soluzioni che stanno attorno alla membrana. A basse concentrazioni la corrente aumenta in modo pressoché lineare con la concentrazione. Per concentrazioni elevate la corrente tende a raggiungere un valore che non aumenta più quando viene aumentata la corrente, a questo punto si dice che la corrente è andata in saturazione. Tale saturazione è in accordo con l’ipotesi che il passaggio di ioni comporti un loro legame con siti particolari del canale stesso, mentre non concorda l’idea che il loro passaggio obbedisca soltanto alle leggi di diffusione elettrochimica, ciò infatti richiederebbe che la corrente continuasse a crescere al crescere della concentrazione.