La fisiologia legata al ritmo sonno veglia è stata studiata fin dai primi anni del 1800. Le prime teorie suggerivano che fosse il risultato di un condizionamento o di una riduzione passiva dell’input sensoriale. Successivamente, la ricerca ha identificato specifiche regioni cerebrali coinvolte nella regolazione del sonno, tra cui la formazione reticolare e l’ipotalamo. Anche il talamo è importante nella regolazione del sonno, con diversi nuclei coinvolti in diverse fasi del sonno. Tuttavia, i meccanismi esatti dell’inizio del sonno e della transizione tra le fasi non sono ancora completamente compresi.
Le prime teorie.

Le ricerche sulla fisiologia del sonno hanno inizio nei primi anni dell’800 con Pavlov. Egli considerò il sonno come il risultato di un condizionamento; uno stimolo condizionato ripetuto in modo monotono ad un cane è in grado di produrre uno stato di sonno. Per Pavlov si trattava di un processo d’inibizione interno, che inizia nelle aree corticali e si diffonde a macchia d’olio lungo tutta la neocorteccia fino ad investire le strutture sottocorticali.
Sebbene Pavlov si fosse opposto alle teorie alternative che postulavano l’esistenza di centri induttori del sonno; questi furono in seguito scoperti da Von Economo (1926) che mise in risalto il ruolo dell’ipotalamo e da Nauta (1946) il quale dimostrò come, nell’animale, la sezione dell’ipotalamo posteriore produca una sonnolenza persistente, mentre le sezioni rostrali causavano insonnia (questa ipotesi è stata confermata dalla ricerca recente).
I lavori di Kleitman (1929) invece considerarono il sonno come un fenomeno passivo, infatti, per l’autore il sonno è il risultato di una riduzione delle afferenze sensoriali che mantengono le condizioni di veglia.
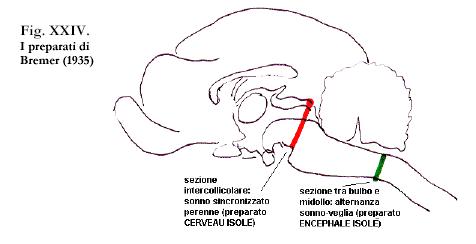
Tale ipotesi sul sonno fu ripresa anche da Bremer (1935). Quest’autore, postulò l’esistenza di un meccanismo regolatore posto nel tronco dell’encefalo.
Le sue ipotesi derivarono dall’osservazione di alcune lesioni sperimentali: il preparato cerveau isolé, in cui isolò chirurgicamente la corteccia dal tronco encefalico, e il preparato encephale isolé in cui la sezione avveniva invece a livello del bulbo. Nella prima situazione l’animale sperimentale presentava uno stato comportamentale ed elettrofisiologico simile all’addormentamento. Nel secondo preparato, invece l’animale manteneva il ritmo sonno-veglia.
Questa fenomenologia fu spiegata da Bremer col fatto che nell’encephale isolé, le afferenze sensoriali, erano in grado di raggiungere la corteccia e consentire all’animale di mantenere lo stato di veglia. Per l’autore, quindi, il sonno era un fenomeno passivo, che s’instaurava attraverso un meccanismo troncoencefalico, in grado di mediare le afferenze sensoriali che giungono alla corteccia.
La formazione reticolare
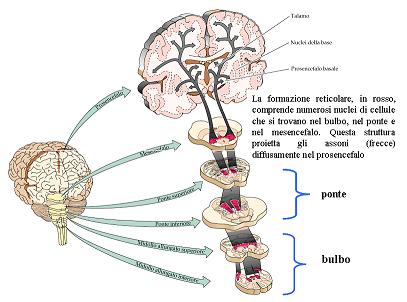
La componente reticolare del tronco, è stata successivamente scoperta da Moruzzi e Magoun(1949). Essi osservarono che la stimolazione elettrica ad alta frequenza della formazione reticolare del tronco, che si estende dal bulbo al mesencefalo, è in grado di produrre risveglio e attivazione corticale (desincronizzazione EEG) nell’animale sperimentale.
Studi successivi (Lindsley e collaboratori, 1950) dimostrarono che la lesione bilaterale del tronco delle vie del lemnisco laterale (quindi una deafferenzazione sensoriale) non impediva il risveglio e l’alternarsi del ritmo sonno veglia. Dall’altra parte invece, la sola lesione della componente reticolare, anche risparmiando le vie sensoriali, produceva il quadro del cervau isolé.
In quegli anni perciò la teoria reticolare, sostituì la teoria di Bremer, in quanto è l’interruzione delle vie reticolo – corticali la vera responsabile dello stato di sonno.
Il sonno: fenomeno attivo vs fenomeno passivo
Ancora il sonno, però, era considerato come un fenomeno passivo causato da una riduzione di attività dei neuroni reticolari ascendenti che produceva la sincronizzazione EEG. Fino agli anni 50, infatti, si pensava che la formazione reticolare avesse una funzione ascendente risvegliante, capace di attivare la corteccia e mantenere i centri ipnogeni del talamo inibiti.
Alla fine degli anni 50 Batini, mise in luce l’esistenza di possibili differenziazioni funzionali all’interno della struttura reticolare. Nel preparato mediopontino (sezione del tronco a livello pontino all’ingresso del quinto paio di nervi cranici), l’autore osservò una insonnia caratterizzata da desincronizzazione EEG e movimenti oculari. Se la sezione era invece eseguita pochi millimetri più rostralmente, l’EEG diventava sincrono e i movimenti oculari scomparivano. Quindi le strutture caudali (ipnogene) inducono sincronizzazione EEG e sonno comportamentale mentre le strutture rostropontine e mesencefaliche sono responsabili della veglia (sono risveglianti) (Mancia, 1996).
Da questo punto in poi predominerà una visione del sonno come un fenomeno attivo determinato daIl’attivitò di numerosi centri nervosi. Iniziano perciò ad essere studiate le singole componenti fisiologiche del sonno:
Meccanismi omeostatici del tronco.
Bonvallet e Bloch (1961) hanno ipotizzato un controllo omeostatico delle strutture bulbari sincronizzanti, le quali hanno un azione inibitoria sui centri ponto-mesencefalici desincronizzanti. Nei loro studi, hanno dimostrato che la stimolazione mesencefalica, provoca un risveglio elettroencefalico. Tale risveglio, aumenta in durata se è seguita una sezione che separa le strutture caudali ipnogene da quelle rostrali risveglianti. Mancia e collaborati (1974) hanno corroborato tale ipotesi. Essi hanno evidenziando che le influenze sinaptiche in direzione rostrocaudale sono essenzialmente di natura eccitatoria, viceversa quelle ascendenti, caudorostrale, sono inibitorie. Quindi i circuiti del tronco sembrano essere mediati da un meccanismo a feed-back negativo che autoregola l’attivazione mesencefalica limitando un possibile eccesso di veglia.
Aree basali preottiche
Nel 1962 Sterman e Clemente hanno osservato che la stimolazione elettrica delle aree basali preottiche era in grado d’indurre sincronizzazione EEG nei gatti. In queste aree sono presenti neuroni GABAergici (Gritti et al., 1993) che proiettano sia alla corteccia sia al nucleo mediodorsale del talamo. La stimolazione elettrica dell’area prosencefalica basale, infatti, produce una oscillazione spontanea eccitatoria/inibitoria dei neuroni mediodorsali del talamo (Mariotti et al., 1994). Perciò, sembra probabile, che i neuroni GABAergici partecipino ai meccanismi sincronizzanti del sonno, sia direttamente attraverso le proiezioni alla corteccia, sia indirettamente, attraverso i nuclei del talamo o le regioni sincronizzanti bulbopontine (Mancia et al., 1976)
Le proiezioni colinergiche delle aree preottiche basali (nucleo di Meynert) sono invece implicate nel meccanismo del risveglio e nel controllo della desincronizzazione del sonno REM (Szymusiak e McGinty, 1990)
Le aree preottiche basali, inoltre costituiscono, un sistema d’interazione tra i meccanismi termoregolatori e ipnogeni. Il riscaldamento di queste aree produce, infatti, sincronizzazione EEG ed addormentamento. Ciò favorisce l’ipotesi che l’induzione dello stato ipnico è facilitato, quando la temperatura ipotalamica supera un certo livello (McGinty e Szymusiak, 1990). Inoltre, poiché nel sonno sincrono si rileva una diminuzione della temperatura, è possibile che questo stato sia un comportamento termoregolativo.(Mancia, 1996 pag 29).
L’ipotalamo posteriore
L’ipotalamo posteriore partecipa assieme alla formazione reticolare mesencefalica ai meccanismi di veglia ed alla regolazione del sonno REM. La stimolazione elettrica di quest’area, infatti, induce desincronizzazione EEG e risveglio comportamentale, mentre la sua deattivazione da raffreddamento induce sincronizzazione e sonno (Naquet et al., 1965). Inoltre, nei preparati cerveau isolé, la stimolazione delle strutture ipotalamiche anteriori e posteriori induce desincronizzazione EEG (Belardetti et al., 1977), suggerendo un ruolo diretto dell’ipotalamo nei processi del risveglio.
Il talamo
Il talamo è una componente fondamentale del sonno, in quanto sottostà ai processi di sincronizzazione e desincronizzazione corticale, ed è la prima stazione di controllo dei segnali afferenti verso la corteccia.
La ricerca sperimentale sul gatto ha messo in luce che i principali nuclei del talamo che sono in relazione con l’inizio ed il mantenimento del sonno sono il nucleo mediodorsale (Marini et al., 1990), il nucleo centralis lateralis (Marini et al., 1992) che invece sembra sottostare alla regolazione della veglia e del sonno REM, ed il nucleo reticolare del talamo (NRT). Steriade e collaboratori (1993) hanno ipotizzato che l’NRT sia fondamentale nei processi sincronizzazione e del sonno comportamentale.
Conclusioni
Da quanto esposto, risulta che i processi di veglia e sonno sono regolati da componenti fisiologiche complesse. Infatti, ancora non è chiaro il meccanismo preciso con cui si realizza l’addormentamento. Una ipotesi (Mancia, 1996) attribuisce l’inizio del sonno all’attivazione dei neuroni (forse GABAergici) delle regioni ipotalamiche anteriori e prosencefaliche. Queste regioni, mettono in moto le sequenze sincronizzanti talamo-corticali e contemporaneamente sincronizzano i neuroni del tronco che così entrano in fase con le sequenze talamiche. Il passaggio dal sonno sincrono alla veglia (o al sonno REM), invece, può essere provocato dall’attivazione dei neuroni ponto-mesencefalici e ipotalamici posteriori, che a loro volta rompono le sequenze talamiche, operando sui neuroni GABAergici del NRT.