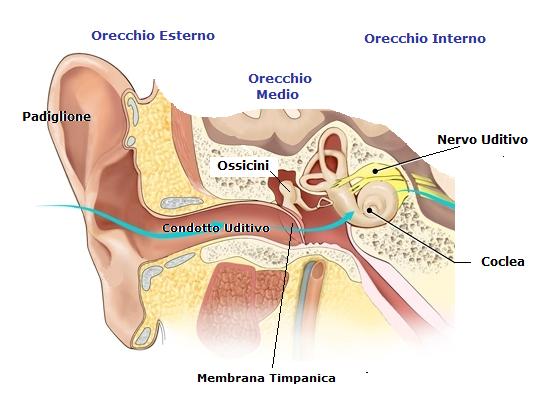
Il sistema uditivo è costituito da diverse strutture anatomiche e neurali che trasducono e trasmettono il segnale acustico. L’orecchio esterno raccoglie le onde sonore e le trasmette all’orecchio medio, dove la membrana timpanica e gli ossicini amplificano le vibrazioni. L’orecchio interno contiene la coclea, un organo a forma di chiocciola che converte le vibrazioni meccaniche in impulsi nervosi. Nella coclea si trova l’organo del Corti, formato da cellule ciliate dotate di stereociglia che si flettono in risposta al movimento della membrana basilare e della membrana tettoria. Questa flessione provoca l’apertura di canali ionici che depolarizzano le cellule ciliate e inducono il rilascio di neurotrasmettitori verso le fibre del nervo acustico. Il nervo acustico porta il segnale nervoso al nucleo cocleare, situato nel tronco encefalico, dove si verifica una prima elaborazione delle informazioni sonore. Da qui il segnale prosegue verso il collicolo inferiore, il corpo genicolato mediale e infine la corteccia uditiva primaria, dove si realizza una mappa tonotopica delle frequenze udibili.
Struttura del sisitema uditivo
Il sistema uditivo converte le vibrazioni prodotte dalle onde sonore in un segnale nervoso ricco di numerose informazioni.
Il sistema uditivo è costituito dall’orecchio esterno, l’orecchio medio e l’orecchio interno. Il compito dell’orecchio esterno è quella di incanalare le onde sonore verso la membrana timpanica. Le pertubazioni prodotte dal suono si irradiano, allontanandosi dalla sorgente che le produce, come onde circolari nelle quali picchi di aumento della pressione nell’aria si alternano con minimi si rarefazione dell’aria stessa. La frequenza dell’onda viene misurata dal numero dei picchi e determina la tonalità. La frequenza viene misurata in cicli al secondo o Hz. L’orecchio umano è sensibile ad una gamma di frequenze acustiche da 20 a 20.000 Hz. L’ampiezza del suono è la massima escursione della pressione dell’aria prodotta da un suono nei due sensi ed è correlata con l’intensità del suono stesso. Essa è misurata in dB (decibel).
Orecchio medio
I suoni che raggiungono l’orecchio viaggiano attraverso il canale acustico esterno o meato e raggiungono l’orecchio medio dove fanno vibrare la membrana timpanica. Tali vibrazioni attraversano l’orecchio medio e sono convogliate da una serie di tre piccoli ossicini, uno dei quali il martello è aderente alla membrana timpanica.
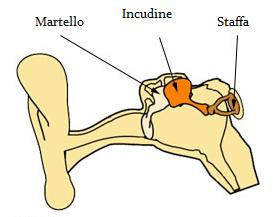
Le trasmissioni del martello vengono trasmesse ad un’apertura della coclea chiamata finestra ovale per mezzo di altri due ossicini l’incudine e la staffa.
Se l’orecchio medio non esistesse, i suoni raggiungerebbero direttamente il liquido della coclea a livello della finestra ovale, e la maggior parte dell’energia sonora verrebbe riflessa, in quanto, i liquidi hanno un’impedenza acustica molto maggiore di quella dell’aria. La superficie della membrana timpanica è maggiore di quella della finestra ovale. Perciò, la pressione totale che agisce su quest’ultima, che è molto più piccola, aumenta notevolmente. Questo deriva dal fatto che gli ossicini agiscono come un sistema di leve in grado di aumentare la pressione sulla finestra ovale.
La coclea
La coclea si avvolge a spirale per due giri e mezzo intorno a un pilastro centrale detto modiolo. E’ suddivisa in tre compartimenti o scale piene di liquido:
- la scala timpanica che segue i contorni esterni della coclea;
- la scala vestibolare, segue i contorni interni della coclea ed è in rapporto con la scala timpanica a livello dell’alicotrema;
- la scala media che s’introflette come un guanto nel canale spirale.
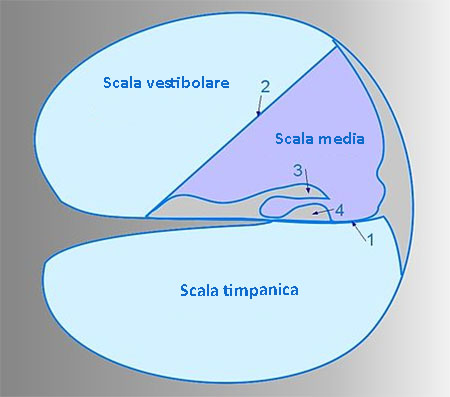
Il suono che entra nell’orecchio fa oscillare la staffa e le seguenti oscillazioni trasmettono energia a tutti i tre componenti della coclea. La staffa oscillando alternativamente verso l’interno o l’esterno della coclea, esercita pressioni variabili sul liquido presente nella scala vestibolare.
Siccome i liquidi sono incompressibili, le onde pressorie determinano un movimento alternato, verso l’interno o l’esterno della membrana che riveste la finestra rotonda della scala timpanica. Le onde pressorie producono movimenti anche nella scala media e nella membrana basilare. L’organo del Corti che è la struttura deputata alla trasmissione sensoriale, riposta sulla membrana basilare, viene stimolato anch’esso dal movimento del liquido.
I compartimenti della coclea sono disposti in modo da convertire la differenza di pressione che si stabilisce tra scala vestibolare e scala timpanica in movimenti oscillatori della membrana basilare che eccitano o inibiscono le cellule sensoriali dell’organo del Corti.
I suoni possono anche evitare l’orecchio medio e raggiungere direttamente la coclea per conduzione ossea e cioè attraverso le vibrazioni di tutto l’osso temporale.
Il test di Rinne permette di distinguere due principali sordità: la sordità di conduzione determinata da un danno all’orecchio medio e la sordità sensori-neuronale determinata da alterazione della coclea.
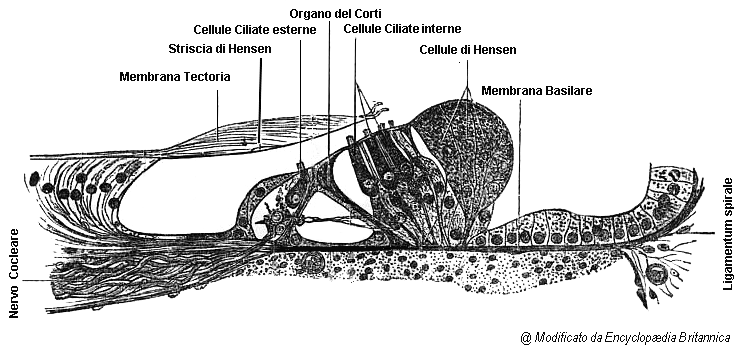
- coclea: struttura anatomica
Orecchio interno
Le cellule recettoriali dell’orecchio interno, le cosiddette cellule ciliate sono localizzate nell’organo del Corti. Vi sono tre file di cellule ciliate esterne e una fila di cellule ciliate interne.
Sulla superficie apicale di ogni cellula ciliata vi è un fascio di stereociglia. Esse contengono una serie di filamenti paralleli di astina legati l’un l’altro che conferiscono loro un certo grado di rigidità.
Le stereociglia vanno a terminare nella sovrastante membrana tettoria. Siccome le estremità delle stereociglia sono impigliate nella membrana tettoria, mentre i corpi delle cellule cigliate riposano sulla membrana basilare, le stereociglia tenderanno a flettersi se la membrana tettoria e la membrana basilare si muovono.
Il movimento delle sterociglia in un senso depolarizza la cellula ciliata provocando l’apertura di canali ionici che determinano, l’insorgenza di una corrente entrante. Il movimento delle stereociglia in senso opposto iperpolarizza invece la cellula stessa. Di conseguenza, quando un suono determina un movimento oscillatorio della membrana basilare, lo spostamento angolare, avanti ed indietro, delle stereociglia provoca l’insorgenza di variazioni sinusoidali di potenziale che hanno la stessa frequenza dello stimolo acustico.
Le cellule ciliate, inoltre, possiedono una serie di strutture specializzate per il rilascio del neurotrasmettitori alla loro estremità basale. A questo livello, infatti, sono a contatto con gli assoni periferici dei neuroni bipolari i cui corpi cellulari sono localizzati nel ganglio spirale e le cui branche danno origine al nervo acustico. Il rilascio del neurotrasmettitore da parte delle cellule ciliate eccita le terminazioni periferiche e ciò determina l’insorgenza di potenziali di azione nelle branche centrali delle cellule gangliari che vanno a costituire il nervo acustico.
Struttura della membrana basilare
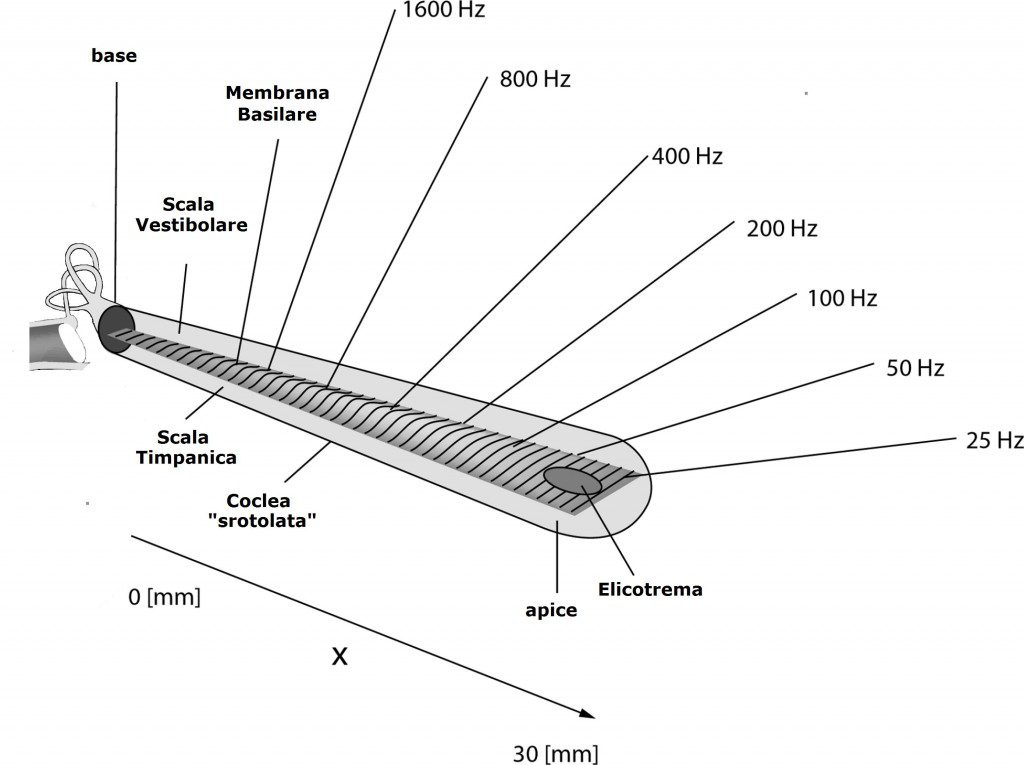
Helmholtz osservò che la membrana basilare possiede delle striature trasversali e che la lunghezza della membrana varia andando dalla base all’apice della coclea. La membrana è rigida e stretta in prossimità della finestra ovale e molto più larga e flessibile all’apice della coclea.
Helmoltz ipotizzò che le striature trasversali delle diverse parti della membrana basilare entrassero in risonanza con frequenze sonore diverse. Le striature vicino alla finestra ovale avrebbero dovuto entrare in risonanza con le frequenze più elevate. Invece, le striature vicino all’apice avrebbero dovuto risuonare con le basse frequenze.
Secondo questa teoria della risonanza, le diverse frequenze acustiche avrebbero dovuto interessare parti diverse della membrana basilare che a loro volta eserciterebbero la propria influenza su popolazioni distinte di cellule ciliate.
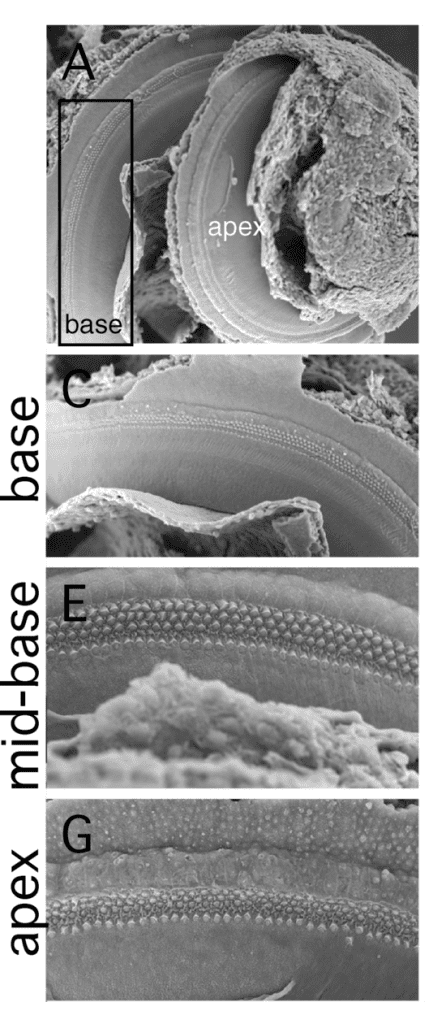
Le cellule ciliate presenti nelle diverse zone della membrana basilare non sono identiche ma differiscono le una dalle altre. La differenze riguardano le loro proprietà elettromeccaniche che potrebbero rappresentare i fattori più importanti nel determinare la loro selettività per le diverse frequenze.
Alla base della coclea, dove la membrana basale è stretta e rigida, le cellule ciliate e le loro stereo ciglia sono corte e rigide. Verso l’apice, le cellule ciliate e le loro stereociglia hanno lunghezza flessibile all’incirca doppie delle cellule situate vicino alla base.
Le onde sonore attiveranno quindi zone diverse della membrana basilare e stimoleranno popolazioni di cellule ciliate diverse, dotate di diversa sintonia meccanica.
Potenziale di riposo nelle cellule ciliate dei vertebrati
Lewis e collaboratori hanno osservato che le cellule di alcuni vertebrati inferiori mostrano oscillazioni spontanee che sono diverse a seconda della loro posizione lungo la membrana basilare.
La stimolazione meccanica depolarizza o iperpolarizza alternativamente la cellula in quanto aumenta o diminuisce l’ampiezza delle oscillazioni di correnti dovute al calcio (Ca) e al potassio (K).
La risonanza elettrica dipende dall’interazione di tre diverse correnti: una di Ca, una di K attivata dai Ca e una corrente K ritardata. La fase di depolarizzazione è dovuta all’ingresso di Ca in prossimità dell’apice della cellula. L’ingresso di Ca attiva anche un canale K sensibile alla concentrazione di Ca dando origine ad una corrente uscente di K. L’effetto iperpolarizzante della corrente uscente K è poi ulteriormente aumentato da una corrente K voltaggio dipendente. Man mano che il potenziale tende a tornare a livelli di riposo, sia la corrente K e Ca attivata, che la corrente K voltaggio dipendente, divengono progressivamente meno intense e la cellula è pronta ad iniziare un altro ciclo. L’interazione delle correnti depolarizzanti con quelle iperpolarizzante determina la comparsa di fluttuazioni spontanee al valore del potenziale di riposo.
Un possibile meccanismo di emissione oto-acustica spontanea
Bromwell e collaboratori hanno esaminato gli effetti di stimoli transcellulari di corrente alternata sulla lunghezza dei corpi cellulari delle cellule ciliate dell’orecchio interno.
I ricercatori hanno scoperto che queste cellule sono in grado di adattare la loro lunghezza in funzione della frequenza e dell’intensità degli stimoli elettrici. Questo fenomeno potrebbe essere correlato al meccanismo dell’emissione oto-acustica spontanea, che consiste nella produzione di suoni dall’orecchio interno senza stimolazione esterna. Si ritiene che in questo caso siano le cellule ciliate a muoversi impartendo uno spostamento dell’endolinfa, che provoca il movimento della staffa e di conseguenza una stimolazione acustica spontanea. Le emissioni oto-acustiche sono considerate un indice della salute dell’orecchio interno e sono generate dalla vibrazione delle cellule ciliate in risposta al suono.
Il ruolo delle cellule ciliate nella trasmissione delle informazioni acustiche
Sensibilità e selettività delle cellule ciliate
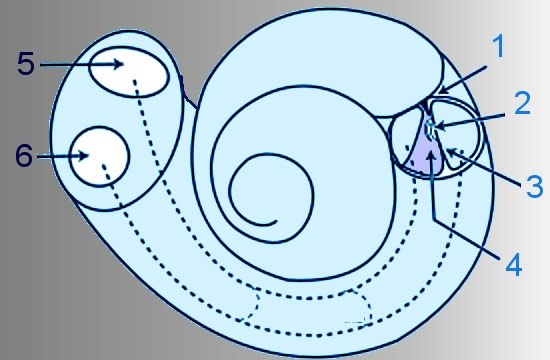
Le cellule ciliate sono innervate dai neuroni bipolari del ganglio spirale localizzato nel modiolo della coclea. Le branche periferiche degli assoni delle cellule del ganglio spirale vengono attivate da un neurotrasmettitore liberato dalle ciliate, mentre le branche centrali degli assoni delle cellule del ganglio vanno a costituire il nervo acustico. Circa il 90% delle fibre di queste cellule innerva le cellule ciliate interne: ogni cellula ciliata interna riceve contatti da circa 10 fibre ed ogni fibra entra in contatto soltanto con una cellula ciliata interna. Il rimanente 10% delle fibre ha un andamento divergente e va a innervare parecchie ciliate esterne (vedi figura: coclea: struttura anatomica).
Cellule ciliate interne ed esterne: differenze strutturali e funzionali
Le cellule ciliate esterne sono innervate da fibre efferenti che provengono dal sistema nervoso centrale, perciò le loro variazioni di lunghezza potrebbero essere sotto il controllo nervoso. Le cellule ciliate interne sono responsabili del riconoscimento dei suoni e trasmettono l’eccitamento alla quasi totalità delle fibre del nervo acustico. La modulazione selettiva delle proprietà delle cellule ciliate esterne può tuttavia modificare significativamente le proprietà meccaniche dell’organo del Corti e fornire in tal modo al sistema nervoso centrale un meccanismo efficiente per sintonizzare l’orecchio verso suoni di particolare interesse.
Inoltre, siccome la maggioranza delle cellule del ganglio innerva una sola cellula ciliata interna, ogni fibra del nervo acustico risponde in modo selettivo ai suoni di frequenza particolare. È infatti possibile descrivere una curva tonale registrando le risposte di singole fibre del nervo acustico in risposta a brevi stimoli sonori di frequenza e di ampiezza diverse.
Le curve tonali sono rappresentazioni grafiche dell’intensità del suono necessaria, per determinare in una fibra del nervo acustico la risposta soglia all’interno della gamma delle frequenze sonore udibili. Le curve tonali sono, perciò, l’espressione della sensibilità della fibra stessa. Sebbene singole fibre siano in grado di rispondere a un vasto ambito di frequenze, ogni fibra risulta, particolarmente selettiva ad una frequenza specifica che viene detta frequenza caratteristica.
L’organizzazione delle vie uditive centrali: nuclei, fibre e sinapsi
Le fibre del nervo acustico terminano nel nucleo cocleare che riposa sulla superficie esterna del peduncolo cerebrale inferiore. Le fibre penetrano attraverso la parte ventrale del nucleo coclearie suddividendone la parte ventrale in un nucleo coclearie antero-ventrale e in un nucleo postero-ventrale. Ogni fibra si suddivide non appena entra nel nucleo coclearie.
Gli assoni delle cellule del nucleo coclearie si suddividono in tre fasci distinti: la stria acustica dorsale, la stria acustica intermedia e il corpo trapezoide.
La via principale è costituita dal corpo trapezoide. Esso contiene le fibre destinate ai nuclei dell’oliva superiore di entrambi i lati del tronco dell’encefalo. Il nucleo mediale dell’oliva superiore è in rapporto con la localizzazione dei suoni in base a differenze interaunali di tempo. Questo nucleo è formato da cellule che ricevono afferenze dai nuclei cocleari controlaterale e ipsilaterali. Il nucleo laterale dell’oliva superiore è principalmente in rapporto con l’analisi delle differenze dell’intensità del suono. Gli assoni del complesso dell’oliva superiore convergono a formare il lemnisco laterale.
In questo modo fin dall’inizio, il sistema nervoso centrale riceve estese afferenze uditive bilaterali. Di conseguenza lesioni delle vie uditive centrali non determinano mai sordità monoaurale. Il lemnisco laterale passa attraverso i nuclei del lemnisco laterale. Anche a questo livello si ha uno scambio di fibre attraverso la commessura di Probst. Tutte le fibre del lemnisco laterale fanno sinapsi infine con le cellule del collicolo inferiore che ricevono afferenze binaurali ed hanno un’organizzazione tonotopica.
La maggior parte delle cellule del collicolo inferiore inviano i loro assoni al corpo genicolato mediale del talamo: queste a loro volta mandano i loro assoni alla corteccia uditiva primaria omolaterale che è localizzata nel giro temporale superiore.
Tonotopia e cellule del nucleo cocleare
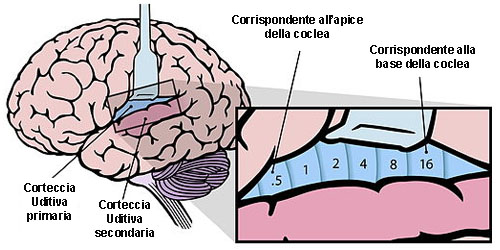
Il nucleo cocleare ha una disposizione topografica che rispecchia l’organizzazione tonotopica delle cellule e delle fibre che lo compongono. Questo è il criterio fisiologico più importante per capire la sua struttura e funzione.
Le fibre acustiche primarie che provengono dalla base della coclea, dove si percepiscono le alte frequenze, penetrano profondamente nel nucleo. Le fibre primarie che provengono dall’apice della coclea, dove si percepiscono le basse frequenze, terminano invece a livelli più superficiali.
Nel nucleo cocleare ventrale si distinguono due tipi principali di cellule: le cellule stellate e quelle a cespuglio. Le cellule stellate, se depolarizzate con uno stimolo di corrente continua, generano una serie di potenziali di azione a intervalli regolari. Questa modalità di scarica è detta risposta alternante e dipende dalla frequenza dello stimolo acustico. Le cellule stellate potrebbero quindi codificare le diverse componenti spettrali del suono. Le cellule a cespuglio, invece, emettono uno o due potenziali di azione all’inizio dello stimolo di corrente continua. Queste cellule potrebbero avere il compito di segnalare l’inizio del suono.
La localizzazione dei suoni: un processo complesso che coinvolge orecchie e cervello
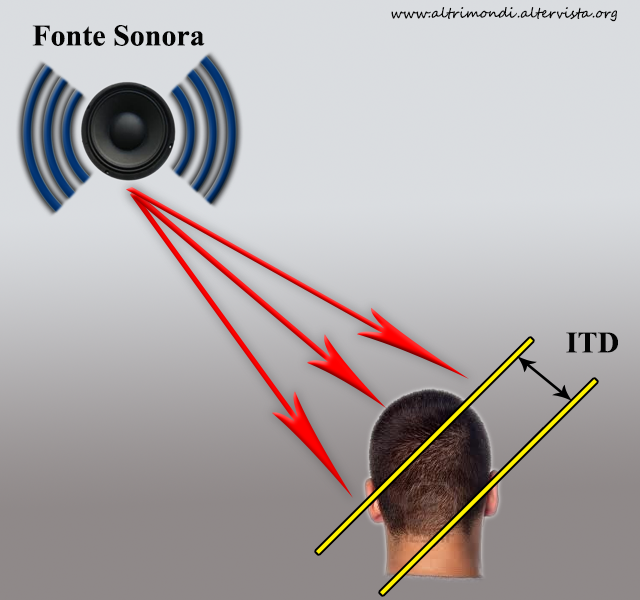
La localizzazione dei suoni è la capacità di riconoscere la direzione e la distanza di una fonte sonora. Questa capacità sfrutta le informazioni provenienti dalle orecchie e i processi cognitivi associati. Si tratta di una facoltà essenziale per la sopravvivenza e l’orientamento degli animali udenti. Questi animali presentano diverse abilità a seconda delle specie; ad esempio, i gufi hanno un’ottima localizzazione dei suoni. Essi, infatti, si basano sull’udito per cacciare le loro prede.
La localizzazione dei suoni è una facoltà che viene realizzata dalle strutture cerebrali paragonando le differenze dell’intensità e il tempo di arrivo dei suoni che arrivano a ciascun orecchio.
Un suono breve come un click, che prende origine da una zona qualunque dello spazio, anzitutto colpirà l’orecchio più vicino. Solo successivamente e dopo un breve ritardo raggiungerà l’orecchio opposto. La durata di questo ritardo è determinata dalla distanza tra le due orecchie, dalla velocità del suono e dalla localizzazione della sorgente del suono.
Se la sorgente del suono è localizzata lungo la linea mediana, sia davanti che dietro la testa, il suono colpirà simultaneamente entrambe le orecchie e il ritardo sarà nullo. Per punti disposti tra questi due estremi esisterà uno spettro continuo di differenze interneuronali del tempo di arrivo dello stimolo (o Interaural Time Difference, ITD).
Questo meccanismo è efficace per i suoni a bassa frequenza, compresi tra 20 e 2000 Hz. Per frequenze più elevate, per le quali la lunghezza d’onda del suono assume valori minori della distanza che separa le due orecchie, le differenze di fase o del tempo di arrivo diventano ambigue. Perciò, il sistema nervoso non è più in grado di distinguere se le differenze di fase riguardano un solo ciclo o più cicli.
Per i suoni ad alta frequenza, superiori a 2000 Hz, il capo funge da schermo verso il suono. In questo modo si crea una differenza di pressione sonora tra le due orecchie chee determina una differenza d’intensità percepita del suono. Questo meccanismo è efficace per i suoni ad alta frequenza, compresi tra 2000 e 20000 Hz.
Mappe tonotopiche nella corteccia uditiva
Anche nella corteccia uditiva primaria, vi sono parecchie mappe tonotopiche che rappresentano l’intero spettro delle frequenze udibili. Il IV strato è quello d’ingresso alla corteccia, mentre il V è collegato al corpo genicolato mediale. Analogamente alle cortecce somatica e visiva, anche la corteccia uditiva è funzionalmente organizzata in colonne. Le cellule hanno per lo più caratteristiche binaurali e tendono a raggrupparsi in due tipi diversi di colonne, a disposizione alternata, dette colonne di sommazione e colonne di soppressione. Nella prima, la risposta binaurale delle cellule è maggiore della risposta monoaurale. Nelle seconde, si osserva la dominanza delle afferenze che provengono da una delle due orecchie.
Infine, la corteccia uditiva possiede anche estensioni callosali. Siccome vi sono estese rappresentazioni delle afferenze di ciascun orecchio in entrambi gli emisferi cerebrali, le lesioni corticali unilaterali non alterano in maniera significativa la percezione delle frequenze acustiche, mentre hanno notevoli conseguenze sulla capacità di localizzare i suoni nello spazio. Ogni emisfero cerebrale è deputato alla localizzazione dei suoni che provengono dal lato controlaterale. Il cervello dell’uomo possiede anche nei lobi frontali e temporali (area di Broca e di Wernicke) aree funzionali in rapporto con la percezione dei suoni del linguaggio.
Gentilissimi,
ho consultato il vostro articolo su “Il sistema uditivo” per la mia tesi di laurea, e avrei bisogno di conoscere il nome dell’autore per poterlo adeguatamente citare.
Ringraziandovi anticipatamente, colgo l’occasione per augurarvi buone feste.
Gentilissimi,
Ho consultato questo articolo per la mia tesi e avrei bisogno di conoscerne l’autore per poterlo citare. Vi ringrazio molto,
Gentilissimi,
Ho consultato questo articolo per la mia tesi e avrei bisogno di conoscerne l’autore.
Grazie mille