L’analisi dell’informazione visiva è allo stesso tempo parallela e gerarchica. Essa è suddivisa in diversi componenti, quali quella per l’elaborazione del colore, del movimento, dell’orientamento, della tessitura della forma e della profondità. Aree corticali distinte sono responsabili dell’analisi specifica e parallela di ciascuna di queste caratteristiche.
L’informazione così codificata è successivamente riunificata nelle aree visive superiori in una singola percezione coerente con ciò che vediamo. La retina ha diversi tipi di cellule ganglionari, e i loro assoni proiettano sul nucleo genicolato laterale (LGN).
Il LGN riceve input da entrambi gli occhi e ha una mappa topografica del campo visivo. La corteccia visiva primaria (V1) riceve input dal LGN. I neuroni in V1 rispondono a stimoli sempre più complessi, e diversi parametri sono analizzati in percorsi paralleli e separati. Il percorso “what” codifica le informazioni sulle caratteristiche dello stimolo, mentre il percorso “where” analizza le informazioni spaziali.
L’anatomia funzionale della corteccia visiva umana è poco conosciuta, ma sembra essere molto complessa tanto quanto quella della scimmia. Man mano che ci spostiamo all’interno del sistema visivo, dalla retina al nucleo genicolato laterale (LGN), alle aree corticali successive, i neuroni visivi rispondono a stimoli sempre più complessi. Diversi parametri dello stimolo sono analizzati in vie parallele e separate.
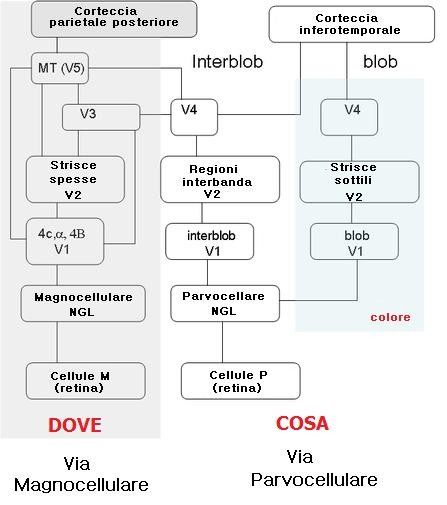
Queste sono divise in due ampi percorsi: la via del cosa (what) e la via del dove (where). La via what codifica le informazioni circa le caratteristiche dello stimolo (come forma e colore) e l’identità degli oggetti, e può essere suddivisa in due ulteriori percorsi colore e forma. La via Where, invece, analizza le informazioni spaziali relative all’oggetto ed è generalmente suddivisa in analisi del movimento e analisi della forma derivante dal movimento.
La retina
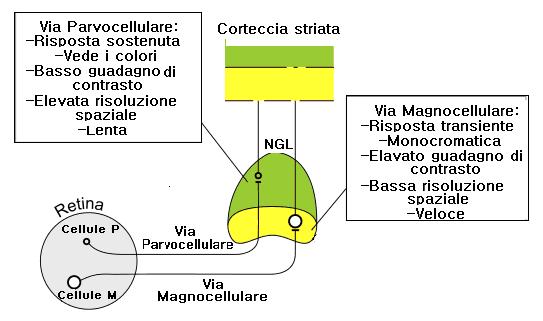
Questa suddivisione inizia ad essere evidente a livello retinico.
La retina dei primati possiede diverse forme di cellule gangliari: due di queste (M e P) costituiscono il 90% di tutte le cellule.
La classe M (cellule A o Pα) costituisce il 10%, mentre il restante 80% è formato dalla classe P (B o Pβ), e i percorsi che si originano da queste cellule sono detti via M e via P.
Le cellule P sono selettive per la lunghezza d’onda e per le alte frequenze spaziali e hanno una risposta lenta e sostenuta (tonica), mentre le cellule M non sono selettive per le lunghezze d’onda, ma lo sono per le basse frequenze ed hanno una risposta fasica e una maggior velocità di conduzione. Il campo dendritico delle cellule M è tre volte più largo di quello delle cellule P.
Il nucleo genicolato laterale (LGN)
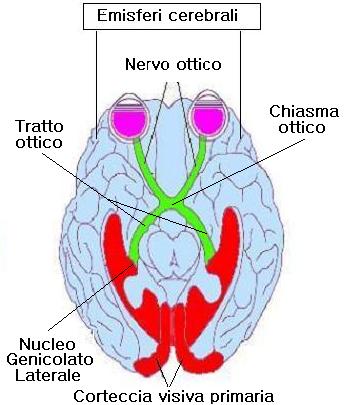
Gli assoni di tutte le cellule gangliari si riuniscono a formare il nervo ottico che fuoriesce dall’occhio e proietta all’LGN dorsale. I nervi dei due occhi si congiungono prima di raggiungere il LGN e formano il chiasma ottico.
Gli assoni delle emiretine nasali s’incrociano e si portano all’altro lato del cervello quindi ogni emisfero cerebrale riceve informazioni dall’emicampo visivo controlaterale.
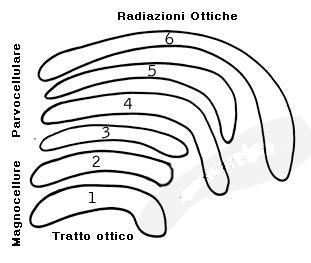
Il LGN consiste in 6 strati, ognuno dei quali riceve segnali in entrata da un solo occhio, gli strati 2,3,5 ricevono informazioni dalle retina ipsilaterale, mentre gli strati 1, 4, 6 dalla retina controlaterale.
La disposizione topografica dei campi recettivi delle cellule gangliari è riprodotta nell’LGN, così ogni strato possiede una mappa dell’emiretina. Gli strati 1 e 2 contengono corpi cellulari più grandi degli altri quattro. I due strati più interni sono perciò denominati strati magnocellulari o M, mentre gli altri esterni parvicellulari o P.
La corteccia visiva primaria (V1)
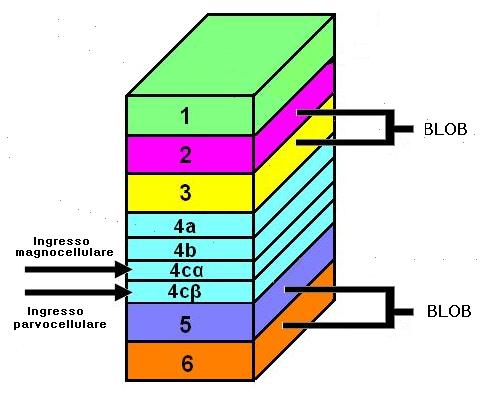
I neuroni dell’LGN proiettano principalmente alla corteccia visiva primaria (corteccia striata o V1). Questa è la prima area visiva corticale e consiste di sei strati principali disposti in bande parallele.
Gli assoni dell’LGN fanno sinapsi con i neuroni dello strato 4. I neuroni degli strati parvocellulari (P) proiettano nella parte più profonda di questo strato (4Cβ), le cui cellule, a loro volta mandano gli assoni agli strati corticali 2 e 3 e da qui a V2. I neuroni degli strati magnocellulari (M) proiettano al sottostrato 4Cα e l’informazione è poi ritrasmessa allo strato 4B e poi ancora a V2 e V5.
Le cellule in 4b sono selettive per l’orientamento e la gran parte di esse mostrano selettività per la direzione del movimento. Alcuni di questi neuroni sono binoculari (rispondono alla stimolazione di entrambi gli occhi) e sono sensibili alla disparità retinica ( differenza relativa degli stimoli nel campo visivo dei due occhi).
Via parvocellulare-blob e via parvocellulare-interblob
La via P si divide e forma due ulteriori nuovi percorsi negli strati superiori di V1. Una via riguarda principalmente il colore ed è denominata via parvocellulare – blob (via P-B). I neuroni della seconda via sono sensibili a caratteristiche quali l’orientamento dello stimolo e sono responsabili della percezione di elevata acuità. Tale percorso è denominato via parvocellulare interblob (via P-I).
Attraverso la colorazione per l’enzima mitocondriale citocromo ossidasi sono stati individuati delle cellule che attraversano gli strati 2 e 3 , 5 e 6. Queste colonne sono denominate blobs. Le aree che circondano i blobs sono dette regioni interblob. Qui la gran parte delle cellule risponde a stimoli con un particolare orientamento, come linee o barre, e ha campi recettivi piccoli. La maggior parte di queste cellule non mostra sensibilità al colore, non mostrano opponenza cromatica e rispondono bene ai bordi di contrasti di luminanza acromatici. I neuroni nelle regioni interblob fanno parte della via P-I.
Le cellule dei blob non sono selettive per l’orientamento ma lo sono per il colore e per la luminosità. Queste cellule sono parte della via P-B. Le cellule a opponenza cromatica dei blob ricevono segnali in entrata dalle cellule P a opponenza cromatica dell’LGN, sebbene nelle cellule blob, il centro del campo recettivo abbia dimensioni maggiori e la codifica del colore ha un’opponenza doppia (per es. il centro potrebbe fornire una risposta ON al verde e di tipo OFF al rosso, mentre la periferia risponderebbe in modo contrario). Per queste ragioni, sembra che i sistemi blob ed interblob lavorino in modi differenti ma complementari.
Aree visiva secondarie
Area visiva 2 (V2)
La destinazione principale delle fibre uscenti da V1 è V2. La colorazione per la citocromo ossidasi non rileva alcuna disposizione dei blobs e regioni interblob, ma presenta un insieme di bande perpendicolari al bordo tra V1 e V2, che si estendono per tutto lo spessore.
Esistono tre tipi di bande: una spessa e l’altra sottile separate da strisce interbanda (strisce pallide). I neuroni dello strato 4B di V1 (parte della via M) proiettano alle strisce spesse. Questi neuroni mostrano le stesse proprietà dei neuroni dello strato 4B (orientamento, movimento e disparità retinica). I neuroni dei blobs proiettano alle strisci sottili, qui le cellule sono selettive per l’orientamento, ma più della metà al colore (doppia opponenza cromatica). Le regioni interblob proiettano alle regioni interbanda i cui neuroni sono selettivi per l’orientamento ma non alla direzione del movimento né al colore. V1 ha un’organizzazione retinotopica, V2 contiene tre mappe. All’interno delle strisce spesse troviamo una mappa per l’orientamento visivo, nelle strisce sottili c’è una mappa per il colore, mentre le regioni interbanda posseggono una mappa per la disparità.
Il percorso M proietta dallo strato 4B di V1 alle strisce spesse di V2. La via P-B proietta dai blobs alle strisce sottili, mentre la via P-I proietta dalle regioni interblob alle aree interbanda.
Area visiva 4 (V4)
Entrambe le suddivisioni della via P, le strisce sottili (colore) e le regioni interbanda (forme) proiettano a V4, e sembra che la separazione tra queste due suddivisioni della via P continui in V4.
Studi di registrazione di singole cellule lasciano supporre che alcune cellule di V4 selettive per il colore, abbiano modalità di risposta più sofisticate rispetto ai neuroni delle aree adiacenti. I neuroni non rispondono alle lunghezze d’onda della luce ma al suo “colore”, un fenomeno conosciuto come costanza del colore.
Lesioni a V4 sembrano compromettere l’abilità delle scimmie di identificare i colori quando variano le condizioni d’illuminazione. Allo stesso modo, lesioni dell’area equivalente negli esseri umani danneggiano la capacità di distinguere i colori, una condizione denominata acromatopsia. E’ interessante notare che lesioni di alcune parti delle aree V1 e V2 negli umani causano la cromapsia, in cui le persone sono in grado di vedere i colori ma non le forme.
V4 è importante anche per la discriminazione degli oggetti, sembra che questi neuroni siano sensibili a semplici forme e oggetti, pertanto lesioni a V4 compromettono la capacità di discriminare gli oggetti. V4 proietta principalmente alla corteccia visiva temporale dove sembra esserci un’integrazione di forme e colore per originare la rappresentazione di oggetti complessi. I neuroni di questa area rispondono a immagini o oggetti complessi, come le facce.
Aree visive 3 e 5 (V3 e V5)
La via M proietta a V3 e V5, sia direttamente tramite lo strato 4b di V1, sia attraverso le strisce spesse di V2.
La maggior parte delle cellule in V3 è selettiva per l’orientamento e si pensa che sia coinvolta nell’analisi delle forme dinamiche. V5 (anche conosciuta come area visiva medio temporale o MT) analizza le informazioni sul movimento e la profondità. Nelle scimmie, lesioni di V5 causano difetti nei movimenti oculari d’inseguimento e nella discriminazione della direzione del movimento.
La via M proietta poi alla corteccia parietale, che sembra essere importante per l’integrazione del movimento e della profondità. Lesioni a questa regione, negli esseri umani, causano una condizione detta sindrome di Balint, che ha tre sintomi principali: primo, difficoltà nel raggiungere gli oggetti (atassia ottica); secondo, aprassia oculare, ovvero difetto della scansione visiva (il soggetto è incapace di mantenere la fissazione degli occhi su un oggetto); terzo, il soggetto è in grado di percepire solo un oggetto alla volta in mezzo a tanti altri (simultanagnosia).
La separazione delle vie P e M non dovrebbe essere troppo enfatizzata; ci sono alcune vie di comunicazione tra i due percorsi. Se gli strati M dell’LGN sono resi inattivi, le risposte dei neuroni risultano ridotte sia in V4, sia nei blob, sia nelle regioni interblob di V1. Ciò suggerisce che il percorso M manda dei segnali in entrata al percorso P.
Lesioni al sistema what (sistema ventrale da V1 ai lobi temporali) rendono difficoltosa la prestazione nei compiti di discriminazione degli oggetti. Lesioni al sistema where (sistema dorsale da V1 ai lobi parietali) pregiudicano le capacità visuospaziali.
I due sistemi poi proiettano a differenti aree corticali prefrontali. Il sistema what proietta alla corteccia della convessità inferiore (IC), mentre il sistema where proietta alla regione dorsolaterale prefrontale (DL). La corteccia prefrontale è una regione importante per la memoria di lavoro; inoltre, esperimenti su animali hanno dimostrato che essa è coinvolta nei processi decisionali.
I due sistemi poi proiettano a differenti aree corticali prefrontali. Il sistema what proietta alla corteccia della convessità inferiore (IC), mentre il sistema where proietta alla regione dorsolaterale prefrontale (DL). La corteccia prefrontale è una regione importante per la memoria di lavoro, inoltre esperimenti su scimmie hanno dimostrato che la regione IC è coinvolta nella memoria di lavoro per gli oggetti, mentre l’area DL è implicata nella memoria di lavoro spaziale.
Si suppone che le vie M e P corrispondano approssimativamente a questi due sistemi, sebbene entrambi sono in comunicazione a tutti i livelli.
Via del “what versus how”
Goodale e Milner hanno proposto un approccio alternativo all’organizzazione what versus where che è stato denominato what versus how.
Una paziente affetta di sindrome di Balint, infatti, non presentava difficoltà a riconoscere il disegno di oggetti comuni, ma non era in grado di afferrare gli oggetti che riconosceva. Ciò fece pensare che lesioni al lobo parietale pregiudichino l’abilità del paziente di usare le informazioni relative alle dimensioni, forma e orientamento di un oggetto al fine di controllare le mani e le dita nell’effettuare il movimento di prensione.
Un’altra paziente che aveva sviluppato un’agnosia per le forme pur avendo notevoli difficoltà nel riconoscere la forma, dimensioni e orientamento di un oggetto mostrava accuratezza nei movimenti delle mani. Quindi nonostante la compromissione delle capacità di discriminazione visiva conscia degli oggetti, le informazioni visive, elaborate inconsciamente, sono state utilizzate dal sistema di controllo delle azioni per dirigere i movimenti di prensione. Ciononostante, la percezione inconscia, può ancora essere possibile in questa condizione (lesioni parietali).
Castello e colleghi hanno descritto una paziente affetta da simultaneoagnosia che era in grado di rispondere alla presenza di oggetti legati tra loro semanticamente, ma non era capace di vedere entrambi gli oggetti. Questi risultati suggeriscono che le pazienti possono essere influenzate da un’inconscia percezione degli oggetti e delle relazioni tra di essi.
Lesioni di V1 causano buchi (scotomi) nel nostro campo visivo. Alcuni pazienti, però, riescono a rispondere a stimoli visivi presentati nei loro scotomi se si richiede loro di effettuare una scelta forzata per indicare i parametri dello stimolo Ad es. un paziente con cecità corticale completa era in grado di seguire con gli occhi una larga immagine a strisce in movimento, sebbene negasse qualsiasi sensazione visiva che potesse spiegare il movimento d’inseguimento dei suoi occhi. Questa percezione inconscia degli stimoli è detta visione cieca.
La retina proietta principalmente al LGN (nucleo genicolato laterale) per mezzo di più di un milione di fibre per occhio, ma proietta anche ad altre strutture come il collicolo superiore (SC). Molte di queste proiezioni trasmettono informazioni circa la posizione, la dimensione, e il movimento degli stimoli visivi. Si crede che queste connessioni possano mediare la visione residua osservata nei soggetti con visione cieca, infatti lesioni dei SC e del pretetto laterale riducono drasticamente la capacità di visione cieca delle scimmie che hanno subito la rimozione di V1.
Una critica leggermente differente, propone la possibile esistenza di una via veloce che oltrepassa V1 e connette direttamente V5 con il LGN e probabilmente anche la retina. Le informazioni che viaggiano lungo questo percorso sembrano essere sufficienti a mediare una grossolana e conscia visione del movimento, e permetterebbero all’individuo di rispondere al movimento degli stimoli più velocemente di quanto accadrebbe se l’informazione giungesse via V1. Questa ipotesi riceve supporto da uno studio di potenziali evocati secondo cui alcune forme di stimoli in movimento attiverebbero V5 prima di V1.