Il potenziale di membrana a riposo
In tutti i neuroni è presente una separazione di cariche elettriche positive e negative disposte sulla superficie interna ed esterna della membrana. A riposo ciò determina il formarsi di un eccedenza di cariche positive sulla faccia esterna della membrana ed un eccesso di quelle negative su quella interna. Questa separazione si mantiene costante e da origine ad una differenza di potenziale elettrico ai capi della membrana conosciuto come potenziale di membrana a riposo.
Quando si verifica un flusso netto di ioni verso l’interno o l’esterno della cellula, la separazione di cariche, esistente ai capi della membrana si modifica. La variazione può determinare un potenziale di membrana meno negativo e quindi una depolarizzazione, o al contrario determinare un potenziale di membrana più positivo e quindi una iperpolarizzazione.
Le risposte passive del potenziale di membrana che non determinano l’apertura dei canali ionici ad accesso variabile vengono dette potenziali elettrotonici, ma quando una depolarizzazione raggiunge un livello critico, a cui si da il nome di soglia, la cellula risponde in modo attivo, con l’apertura dei canali voltaggio dipendente e innesca un potenziale di azione.
Il potenziale di equilibrio per il K
Nelle cellule gliali gli ioni di sodio (Na+) e di cloro (Cl-) sono maggiormente concentrati all’esterno della cellula, mentre quelli del potassio (K) sono più concentrati all’interno, poiché le membrane di queste cellule possiedono canali passivi che sono permeabili soltanto agli ioni K.
Poiché gli ioni K sono presenti in concentrazioni elevate all’interno della cellula essi tenderanno a diffondersi fuori dalla cellula seguendo il proprio gradiente di concentrazione. Il risultato sarà la presenza di un eccesso di cariche positive all’esterno della cellula ed un eccesso di cariche negative all’interno ( a causa della diminuzione di K e della prevalenza di anioni).
Questo processo di diffusione del K da origine ad una differenza di potenziale: positiva all’esterno e negativa all’interno. A seguito di questa differenziazione di potenziale si stabilisce una forza elettrica che si oppone a quella del gradiente di concentrazione ed impedisce agli ioni K di continuare a fluire.
Raggiunto un certo potenziale la carica elettrica diviene uguale a quella chimica (gradiente di concentrazione) e di conseguenza il flusso di K si arresta. Questo avviene quando è stato raggiunto il potenziale di equilibrio per il K (Ek), il movimento dei K verso l’esterno, promosso dal gradiente di concentrazione, è equilibrato da un flusso di K verso l’interno determinato dalla differenza di potenziale.
La permeabilità delle cellule nervose agli ioni K, Na e Cl
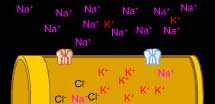
A differenza delle cellule gliali, le cellule nervose sono permeabili a riposo oltre che agli ioni K anche al Na e al Cl.
Gli ioni Na sono maggiormente concentrati all’esterno della cellula e tendono quindi ad entrare all’interno. Inoltre, il potenziale elettrico negativo all’interno della cellula sospinge anche gli ioni Na verso l’interno.
L’ingresso di Na depolarizza la cellula, ma il numero di canali permeabili al K è maggiore di quelli per il Na. Ciò significa che il potenziale di membrana si sposterà solo leggermente al di sopra del valore di equilibrio del K.
Quando il potenziale di membrana si depolarizza, i flussi di K non saranno più in equilibrio e ciò ne determinerà la fuoriuscita dalla cellula. Questo flusso di K uscente tenderà a controbilanciare l’ingresso di Na. Tanto più la cellula si depolarizza tanto più forte sarà la forza elettromotrice che sospinge i K fuori.
Infine, il potenziale di membrana si arresterà ad un nuovo valore di equilibrio che controbilancerà il flusso di Na entrante con il flusso di K uscente. Questo nuovo valore di equilibrio è generalmente intorno a -60mV.
Quando il potenziale di membrana si depolarizza, i flussi di K non saranno più in equilibrio e ciò ne determinerà la fuoriuscita dalla cellula. Questo flusso di K uscente tenderà a controbilanciare l’ingresso di Na.
La pompa sodio – potassio
Per mantenere il potenziale di membrana di riposo stabile, è necessario che l’ingresso di cariche positive sia bilanciato dall’efflusso di cariche dello stesso segno. Se questi flussi non fossero uguali, la separazione di cariche presenti sulle due facce della membrana varierebbero continuamente.
Questa dissipazione dei gradienti ionici viene impedita da una pompa sodio-potassio che sospinge Na e K contro il loro gradiente elettrochimico. Essa, infatti, estrude 3 Na per ogni 2 K. Questo flusso uscente di cariche positive tende ad iperpolarizzare la membrana. La pompa richiede, però, un apporto di energia sotto forma di ATP che è una proteina integrale di membrana di notevoli dimensioni.
La pompa per il cloro
Se la cellula non possiede una pompa per il Cl, il potenziale di riposo dipenderà soltanto dai flussi di Na e K. In questo caso, la concentrazione intracellulare del Cl potrà variare liberamente, e questi ioni si disporranno ai capi della membrana secondo i loro equilibrio elettrochimico.
Nelle cellule nervose, che, invece, possiedono una pompa per il Cl, il trasporto di questi ioni è diretto verso l’esterno, e il gradiente di concentrazione per i Cl aumenterà rendendo il potenziale d’equilibrio per il Cl più negativo del potenziale a riposo.
Eventi scatenanti il potenziale di azione
Una depolarizzazione sopra soglia determina la rapida apertura di alcuni canali Na voltaggio dipendenti. L’aumento di permeabilità verso il Na fa si che l’ingresso di Na divenga maggiore dell’efflusso di K. Questo flusso di cariche positive nette all’interno della cellula, comporta l’ulteriore depolarizzazione di quest’ultima. L’accresciuta depolarizzazione, provoca l’apertura di altri canali Na voltaggio dipendenti.
Questo ciclo rigenerativo a feed-back positivo sospinge il potenziale della membrana verso il potenziale di equilibrio per il Na (+55mV). Questo potenziale potrebbe mantenersi tale se non intervenissero due meccanismi che tendono a ripolarizzare la membrana, ponendo termine al potenziale di azione.
In primo luogo, la costante depolarizzazione tende ad inattivare i canali Na voltaggio dipendenti. In secondo luogo l’apertura ritardata dei canali K voltaggio dipendente che determina un efflusso di K ritardato che si combina con un diminuito ingresso di Na.
Il modello del circuito equivalente del neurone: come rappresentare le proprietà elettriche della membrana neuronale con elementi elettrici semplici.
Il neurone è una cellula specializzata nella trasmissione dell’impulso nervoso, che dipende dalle proprietà elettriche della sua membrana. Per studiare queste proprietà, si può usare il modello di un circuito equivalente del neurone, che simula il comportamento della membrana con elementi elettrici semplici.
Il circuito è formato da un conduttore, una batteria e un condensatore per ogni tipo di ione che attraversa la membrana. Il conduttore rappresenta la resistenza offerta dalla membrana al flusso ionico, la batteria rappresenta la forza che spinge gli ioni a diffondersi lungo il loro gradiente di concentrazione e di potenziale elettrico, e il condensatore rappresenta la capacità della membrana di separare le cariche elettriche tra i due lati.
Il potenziale di membrana a riposo si calcola sommando i contributi delle singole batterie pesati in base alle resistenze dei conduttori. Il potenziale d’azione si genera quando uno stimolo depolarizza la membrana al di sopra di una soglia critica, modificando le resistenze dei conduttori e facendo variare il potenziale di membrana in modo rapido e reversibile.
Il modello del circuito equivalente del neurone può essere adattato a diversi tipi di neuroni, tenendo conto della presenza o meno della guaina mielinica, che aumenta la resistenza della membrana e diminuisce la sua capacità, rendendo la conduzione dell’impulso più veloce ed efficiente.
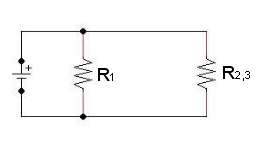
Esempio di un circuito equivalente del neurone
- Il doppio strato lipidico è un cattivo conduttore perché è impermeabile agli ioni, ma ogni canale K può essere rappresentato come un conduttore r, in modo tale che la corrente che passa attraverso i canali K possa essere calcolata attraverso la legge di Ohm (i=V/r).
- Una sorgente di potenziale elettrico viene anche detta forza elettromotrice e una forza elettromotrice generata da una differenza di potenziale chimico viene anche detta batteria. Potremmo perciò rappresentare il potenziale elettrico che si stabilisce ai capi di ciascun canale K come una batteria. Il potenziale sviluppato da una batteria sarà uguale al potenziale di equilibrio per il potassio (Ek) che si aggira intorno a –75mV.
- Ovviamente una cellula possiede molti canali passivi K che possono essere combinati in un circuito equivalente costruito da un conduttore in serie con una batteria. Il valore totale della conduttanza è uguale al prodotto del numero dei canali passivi per la conduttanza. Poiché la batteria di questo circuito è unicamente determinata dal gradiente di concentrazione dell’ione K, il suo valore sarà semplicemente Ek.