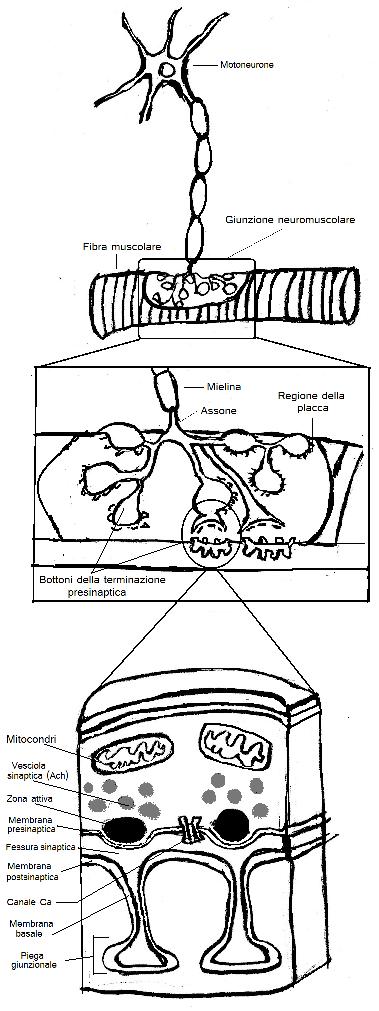
La sinapsi fra il motoneurone e il muscolo scheletrico costituisce l’esempio meglio conosciuto di trasmissione chimica diretta presente nei vertebrati.
Inoltre, la disposizione anatomica delle sinapsi neuromuscolare è piuttosto semplice. Nei vertebrati, ogni fibra motrice innerva una sola fibra muscolare. Il neurotrasmettitore liberato è l’Ach (acetilcolina) e il recettore presente sulla membrana del muscolo è il recettore per l’Ach di tipo nicotinico (ionotropico). L’assone della fibra motrice innerva una zona specifica della membrana muscolare che è detta placca motrice. In prossimità della placca, la fibra motrice perde la guaina mielinica e si suddivide in numerose branche terminali. Ciascun di queste forma alla sua estremità dei bottoni sinaptici al livello del quale viene liberato il neurotrasmettitore. I bottoni sono situati in corrispondenza di depressioni della fibra muscolare dette pieghe giunzionali che contengono i recettori per l’Ach. Su queste pieghe si stende una membrana basale che ricopre tutta la superficie della fibra muscolare.
Ogni bottone presinaptico contiene tutti i componenti necessari per la liberazione del neurotrasmettitore. Infatti, questi elementi comprendono le vescicole sinaptiche che contengono l’Ach e le zone attive. Ogni bottone inoltre contiene canali calcio (Ca) voltaggio dipendenti che promuovono la fusione delle vescicole sinaptiche con la membrana della terminazione.
Le zone attive sono situate di fronte alle pieghe giunzionali della cellula postsinaptica. Nella zona sottostante l’apice delle pieghe la membrana del muscolo è ricca di canali sodio (Na) voltaggio dipendente, che trasformano il potenziale di placca in un potenziale di azione. Il potenziale postsinaptico eccitatorio è chiamato potenziale di placca.
Fatt e Kartz dedussero che il potenziale sinaptico è dovuto ad un flusso di corrente entrante localizzato nella regione della placca motrice, che si propaga passivamente. Il flusso di corrente è però limitato alla regione di placca, in quanto le proteine dei recettori per l’Ach sono localizzate in questa zona.
L’insorgenza rapida del potenziale di placca è dovuta all’aumento improvviso della concentrazione di Ach nella fessura sinaptica. Tuttavia, non tutta l’Ach raggiunge i recettori in quanto si mettono in moto due diversi processi deputati a ridurre la concentrazione di Ach nella fessura sinaptica:
a) l’Ach viene idrolizzato dall’enzima acetilcolinesterasi localizzato nella membrana basale.
b) L’Ach diffonde rapidamente fuori della fessura sinaptica.
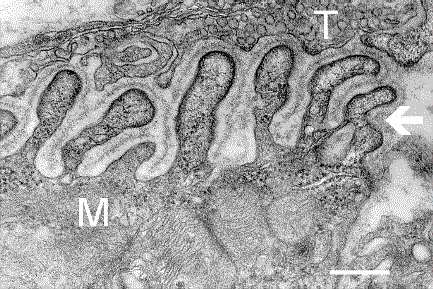
Sono visibile le pieghegiunzionali. Nella figura M indica la fibra muscolare mentre T l’assone terminale
L’andamento temporale e le proprietà dell’andamento del potenziale di placca sono stati analizzati con la tecnica del blocco del voltaggio. Queste ricerche hanno messo in luce che la corrente di placca insorge e decade molto velocemente, ciò può essere dovuto dalla rapida chiusura e apertura dei canali ionici dell’Ach, e dalla costante di tempo passiva.
Il potenziale d’inversione del potenziale di placca è il potenziale di membrana al quale la corrente sinaptica si riduce a zero. Al potenziale d’inversione non vi è alcun flusso netto di corrente in quanto le correnti entranti sono equilibrate da correnti uscenti di egual intensità. Il potenziale d’inversione del potenziale di placca coincide con il potenziale di equilibrio di qualche specie ionica. Se il potenziale di placca fosse dovuto soltanto all’ingresso di Na, il suo potenziale d’inversione sarebbe uguale al potenziale d’equilibrio degli Na (ovvero +55mV). In realtà questo potenziale non è solo dovuto ad una sola specie ionica ma ai flussi di una combinazione di ioni diversi.
La corrente del potenziale di placca dipende da quattro fattori:
- Il numero totale dei canali della placca motrice
- La probabilità di apertura dei singoli canali
- La conduttanza di ogni canale aperto
- La forza elettromotrice che agisce sugli ioni
La probabilità che un canale sia aperto dipende in gran parte dalla concentrazione del neurotrasmettitore a livello del recettore e non dal potenziale di membrana, i canali si aprono per via del loro legame con l’Ach e non per via del loro voltaggio. La fase di crescita rapida della corrente di placca è dovuta all’apertura quasi sincrona di quasi 200.000 canali in risposta al rapido aumento della concentrazione dell’Ach nella fessura sinaptica. La concentrazione di Ach poi diminuisce rapidamente sia per idrolisi che per effusione dell Ach.
Studi biochimici hanno messo in evidenza che il recettore nicotinico comprende 5 subunità disposte in modo da creare un poro centrale. Quando due molecole di Ach si legano alla subunità alfa, il recettore canale cambia conformazione determinando l’apertura di un poro. Sia il Na che il potassio (K) attraversano il canale seguendo il proprio gradiente elettrochimico. Questo canale a differenza dei due diversi tipi di canale voltaggio dipendente, è sufficientemente largo per far passare sia gli ioni Na che i K con selettività approssimativamente uguale.
Una seconda differenza fra i canali voltaggio dipendenti e quelli attivati dai neurotrasmettitori è che il flusso dei Na attraverso canali voltaggio dipendente è rigenerativo. Al contrario, il numero dei canali attivati dall’Ach che si aprono nel corso di un potenziale di azione dipende esclusivamente dalla quantità di Ach. In questo caso la depolarizzazione prodotta dall’ingresso di Na non ha la proprietà di determinare l’apertura di un numero maggiore di canali Ach. Per dare origine a un potenziale di azione, il potenziale di placca deve essere in grado di attivare i canali voltaggio dipendenti circostanti.