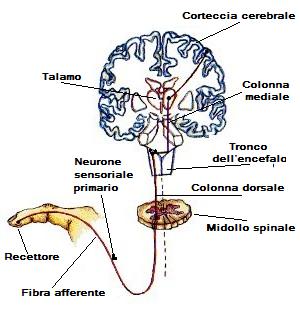
Il sistema somatosensoriale è responsabile della percezione tattile e della posizione delle parti del corpo. Ha recettori nella pelle e nei muscoli che trasmettono segnali nervosi al cervello. Il percorso somatosensoriale comprende tre livelli di neuroni: i primi nel midollo spinale, i secondi nel bulbo e i terzi nel talamo. La destinazione finale dei segnali è la corteccia somatosensitiva primaria nel lobo parietale.
Organizzazione del sistema somatosensoriale
Il tatto ci permette di apprezzare le caratteristiche delle superfici degli oggetti. La propriocezione concerne il senso di posizione statica e dinamica delle dita e degli arti. Mediante il tatto e la propriocezione è possibile rilevare le dimensioni e la forma degli oggetti. Il sistema somatosensoriale è implicato anche nella nocicezione e nella sensibilità termica.
Il sistema somatosensoriale (o somatosensitivo) possiede terminazioni primarie situate nella cute che fungono da recettori e trasformano l’energia degli stimoli in eventi nervosi. Le capacità discriminative tattili sono molto sviluppate all’estremità delle dita.
I recettori dei muscoli, delle dita e delle articolazioni inviano informazioni sulle angolazioni assunte che sono importanti per il riconoscimento degli oggetti.
Nella sistema somatosensoriale , fra i neuroni sensitivi periferici e la corteccia cerebrale sono presenti solo tre sinapsi. La prima è stabilita dal ramo centrale delle cellule dei gangli delle radici dorsali con neuroni del nucleo gracile e cuneato del bulbo. Gli assoni di questi nuclei bulbari attraversano la linea mediana ed ascendono al nucleo ventroposteriore del talamo. Infine, i neuroni talamici inviano i loro assoni ad una regione del lobo parietale anteriore denominata corteccia somatosensitiva primaria o S-I (giro postcentrale).
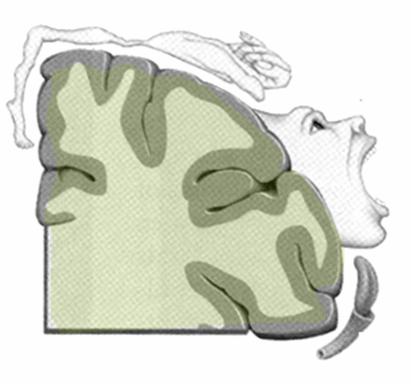
Homunculus: relazione fra organizzazione anatomica del sistema somatosensoriale e specifiche funzioni percettive
Operando su pazienti in anestia locale Penfield stimolava la superficie di S-I in diversi punti e chiedeva ai pazienti cosa sentissero. Egli osservò che la stimolazione di singoli punti del giro postcentrale produceva sensazioni tattili in determinate sedi controlaterali del corpo. Penfield procedendo in questo modo tracciò una mappa della rappresentazione neuronale della corteccia somatosensoriale . Le varie parti del corpo sono rappresentate in aree di dimensioni proporzionali alla loro importanza ai fini della percezione sensitiva.
La superficie del corpo è rappresemtata somatotopicamente a livello della corteccia somatosensoriale. Le regioni somaticamente più importanti nella discriminazione tattile come le dita della mano e la punta della lingua hanno una rappresentazione corticale molto estesa, che è in relazione alle densità d’innervazione (vedi Homunculus).
I disturbi del sistema somatosensitivo possono essere localizzati con notevole accuratezza. Questo perché esiste una relazione diretta fra organizzazione anatomica del sistema nervoso e specifiche funzioni percettive e motorie.
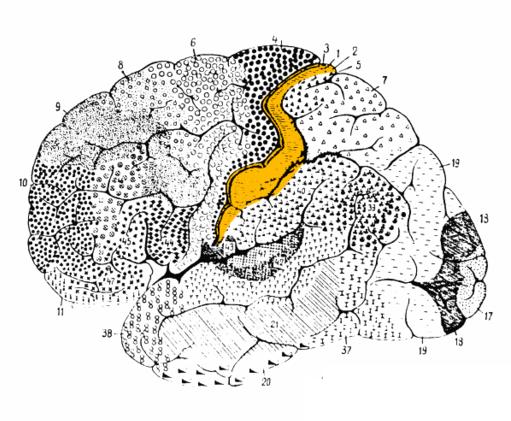
L’introduzione dei microelettrodi ha permesso di dimostrare che al livello della corteccia S-I esistono 4 mappe quasi complete. Ogni mappa è localizzata in una delle 4 aree di Brodmann 3a,3b,1 e 2. Le informazioni sensitive, provenienti dai muscoli e dalle articolazioni, che sono importanti per la propriocezione, sono trasmesse all’area 3a. Le proiezioni provenienti dalla cute raggiungono l’area 3b. Le informazioni sensitive provenienti dalla cute sono ulteriormente analizzate a livello 1 e insieme a quelle provenienti dai muscoli a livello dell’area 2. Di queste quattro rappresentazioni quelle dell’area 3a e 3b sono elementari, quelle delle aree 1 e 2 sono più complesse.
S-I proietta ad altre aree del lobo parietale, ad esempio,alle aree somatosensitive secondarie. A questo livello le informazioni sono utilizzate per l’apprendimento di nuove discriminazioni sensoriali. Le proiezioni alla corteccia parietale posteriore, sono utilizzate per la costruzione dell’immagine corporea.
Principi di funzionamento neurale delle mappe corticali
Le mappe corticali variano da individuo ad individuo in rapporto al loro uso. A questo proposito, in alcune ricerche, una scimma è stata indotta ad utilizzare solo il dito medio per far ruotare un disco per ottenere cibo come compensa. Dopo molte ruotazioni l’area della corteccia devoluta al dito medio risultava molto espansa, a spese della rappresentazione delle dita adiacenti.
Perciò, la pratica rafforza ed espande la rappresentazione corticale delle dita che sono utilizzate di più. In un altro studio, parecchie scimmie sono state studiate a distanza di 10 anni dopo la deafferentazione completa dell’arto superiore. In questi studi, la deafferentazione dell’arto è stata ottenuta mediante sezione di tutti i nervi sensitivi. Negli animali studiati, la rappresentazione della faccia, che è adiacente a quella dell’arto superiore, è risultata espansa. Infatti, si era estesa nell’aria che in precedenza era occupata dalla rappresentazione della mano.
Risultati sperimentali recenti, indicano che nel sistema somatosensoriale le connessioni afferenti ai neuroni corticali si formano in base alla presenza di scariche neuronali correlate tra di loro. Cellule che scaricano in sincronia si connettono tra di loro. Verificando questa ipotesi, mediante intervento chirurgico, sono state unite due dita adacenti in una scimma. In questo modo, le due dita saranno sempre usate insieme. L’aumento di correlazione dell’attività delle due dita determina l’abolizione della netta discontinuità che di norma caraterizza le aree corticali che ricevono afferenza da queste dita.
La sindrome dell’arto fantasma e la rimappatura corticale dell’area somatosensoriale.
Molti soggetti amputati continuano a sentire l’arto mancante e addirittura avvertono dolore provenire da questo. A tale fenomeno è stato dato il nome di sindrome dell’arto fantasma. Gli studi su questi soggetti suggeriscono che le vie afferenti, che di norma raggiungono aree corticali adiacenti alla rappresentazione della mano (faccia spalla), si espandono nell’area precedentemente occupata dalle afferenze provenienti dalla mano. Questo fenomeno è stato detto rimappatura delle sensazioni riferite. In questi pazienti la stimolazione tattile di un’area della faccia o del braccio faceva insorgere sensazioni che venivano riferite alla mano amputata. Questo fenomeno si verifica in quanto l’area della mano nella corteccia è disposta in vicinanza di quella della faccia e del braccio, e perciò può venir controllata dalle afferenze che raggiungono queste aree.
Suddivisione dell’ informazione somatosensoriale in moduli funzionali elementari.
La regione della superficie cutanea che eccita una cellula nervosa è definita campo recettivo. Ogni punto della cute viene monitorato da una popolazione di cellule in quanto i campi recettivi dei vari neuroni sono tra loro sovrapposti.
La maggior parte delle cellule nervose di S-I media una sola modalità: tattile, pressoria, termica o dolorifica. A livello delle varie stazioni sinaptiche del sistemia somatosensitivo, le cellule che rispondono agli stessi stimoli tendono ad essere raggrupate.
Un esempio di questa proprietà fu fornita da Mountcastle. Egli scoprì che i neuroni di tutti e sei gli strati cellulari di una colonna o striscia della corteccia, che va dalla superficie corticale alla sostanza bianca, sono attivati da una sola classe di recettori. Tutti i neuroni di una colonna inoltre ricevono afferenze dalla medesima zona cutanea. I neuroni di una colonna quindi costituiscono un modulo funzionale elementare della corteccia.
Organizzazione e integrazione delle informazioni somatosensoriali
L’integrazione delle informazioni provenienti dallo spazio personale (la superficie del corpo) e da quello peripersonale (lo spazio relativo agli oggetti posti ad una distanza tale da non poter essere raggiunti con un semplice movimento del braccio), si basa essenzialmente su 3 principi:
- Le informazioni relative a stimoli diversi convergono a cellule comuni
- Le proprietà di risposta dei neuroni appartenenti a livelli successivi di elaborazione dell’informazione diventano via via più complesse.
- Le dimensioni dei campi recettivi diventano via via più grandi ad ogni livello di elaborazione
Le aree 3a e 3b di Brodmann costituiscono i primi livelli di elaborazione corticale. Al livello 1 e 2 di queste aree sono presenti neuroni in grado rilevare la direzione e l’orientamento di uno stimolo. Infatti, la 1 e la 2 sono aree implicate nella percezione tattile della forma tridimensionale (sterognosia) e nella discriminazione del movimento.
Le aree 3a e 3b ricevono informazioni dal talamo, mentre le aree 1 e 2 ricevono informazioni convergenti dalle aree 3a e 3b. In tal modo sono in grado di operare il riconoscimento della forma tridimensionale degli oggetti, e di guidare l’esecuzione dei movimenti fini (l’area 2 sembra inviare afferenze alla corteccia motoria primaria).
Corteccia parietale posteriore
Le area somatosensoriale primaria e secondaria della corteccia parietale anteriore, proiettono alle principali suddivisioni del lobo parietale: la corteccia parietale posteriore (aree 5 e 7 di Brodmann). Queste aree ricevono afferenze anche dai sitemi visivi ed uditivi in modo tale da guidare l’attenzione verso lo spazio extrapersonale.
Lesione della corteccia parietale posteriore
Lesioni di queste aree provocano deficit complessi e comprendono deficit di percezione spaziale dell’integrazione visuol-motoria dell’attenzione. Lesioni al lobo parietale posteriore possono determinare afasie ed anche agnosie (incapacità di elaborare percezione degli oggetti).
Una forma di agnosia che compare a seguiti di lesione della corteccia parietale posteriore è l’astereognosia. Essa consiste nell’incapacità di riconoscere col tatto la forma degli oggetti.
Lesioni del lobo parietale posteriore non dominante non provocano disturbi del linguaggio, al contrario pazienti con lesioni in queste aree presentano sorprendenti deficit cognitivi. Essi spesso presentano una forma di agnosia caratterizzata da un peculiare deficit dell’immagine corporea e della percezione delle relazioni spaziali. Infatti, questi pazienti hanno perso la cognizione di tutte le informazioni provenienti dal lato sinistro del loro corpo e dello spazio esterno. Essi ignorano metà del loro corpo (sindrome di negligenza) e non curano di vestire e lavare quella parte del loro corpo. Questi pazienti arrivano perfino a negare l’esistenza del braccio o della gamba. Inoltre, possono ignora54 non solo la metà del loro corpo, ma anche la metà del mondo esterno. E’ presente anche una perdita di memoria e questo fatto spiega perché non siano in generale consapevoli del loro deficit.
Corteccia parietale posteriore e processi attentivi.
Ricerche condotte su soggetti normali hanno dimostrato, che quando l’attenzione verso oggetti nel campo visivo viene cambiata, si attiva la corteccia parietale superiore e quella frontale. La regione parietale viene attivata quando l’attenzione viene cambiata in relazione a stimoli sensoriali. La regione frontale, invece diventa attiva solo quando i segnali-stimolo determinano una risposta motoria.
Quando a pazienti con lesioni del lobo parietale sono presentati simultaneamente due stimoli, uno del campo visivo sinistro e l’altro nela campo visivo destro, essi non si accorgono dello stimolo presentato dalla parte controlaterale alla lesione. Questo deficit a cui è stato dato il nome di estinzione sembra essere la conseguenza dell’incapacità di spostare l’attenzione nella direzione dello spazio controlaterale alla lesione.
Il ruolo della corteccia parietale posteriore nei processi attentivi è stato studiato anche a livello cellulare. Le risposte dei neuroni di quest’area ad una serie di stimoli identici non sono uniformi, ma variano nettamente a seconda dell’attenzione del soggetto. Le cellule forniscono una risposta più intensa quando l’animale è in attesa dello stimolo e meno intensa quando lo ignora. Queste indagini suggeriscono che quando un soggetto dirige lo sguardo verso un oggetto, le cellule della corteccia parietale posteriore che rispondono ad esso, iniziano a scaricare vivacemente. Man mano che il soggetto muove gli occhi per esaminare ulteriormente l’oggetto, anche cellule di altre aree cerebrali implicate nella coordinazione visuomotoria tendono a scaricare maggiormente. Perciò l’attenzione aumenta selettivamente l’attività delle cellule di regioni cerebrali che prendono parte all’elaborazione delle informazioni selezionate.