La trasduzione visiva è il processo mediante il quale la luce viene convertita in segnali elettrici nella retina. Questo processo coinvolge i fotorecettori (coni e bastoncelli), che hanno caratteristiche strutturali e funzionali diverse. I coni sono responsabili della visione diurna e dei colori, mentre i bastoncelli consentono la visione notturna e monocromatica. I fotorecettori contengono i fotopigmenti (rodopsina e iodopsine), che sono formati da una parte proteica (opsina) e da un cromoforo (retinale). Quando il retinale assorbe un fotone, cambia conformazione e attiva una cascata intracellulare che porta alla chiusura dei canali cationici e all’iperpolarizzazione dei recettori. Il calcio ha un ruolo di retroazione e di adattamento alla luce, modulando l’attività della fosfodiesterasi, della guanilato ciclasi e dei canali cationici. Successivamente, le informazioni visive sono trasmesse al cervello attraverso le cellule gangliari, che hanno campi recettivi che rispondono alle differenze di luminosità locale. Un esempio di campo recettivo è quello formato da una zona centrale circondata da un anello di segno opposto: se la zona centrale è eccitata dalla luce, l’anello è inibito e viceversa.
I coni e i bastoncelli

Dopo la focalizzazione sulla retina dell’immagine, essa deve essere trasformata in un’attività neuronale. Questa trasformazione prende il nome di trasduzione visiva ed è attuata dalle cellule recettrici presenti nella retina. Esistono due tipi di fotorecettori: i bastoncelli e i coni. La retina umana contiene circa 120 milioni di bastoncelli e 6 milioni di coni. I coni sono concentrati in una piccola area denominata fovea e sono responsabili alla visione diurna e permettono la visione dei colori. I bastoncelli invece consentono la visione notturna, monocromatica e di bassa acuità. Ci sono tre tipi di coni: 5-10% sono sensibili al blu e formano un anello intorno al bordo della fovea. Il resto è costituito da coni sensibili al rosso e al verde in proporzione di 2:1. queste due ultime due classi sono casualmente mescolate a formare dei piccoli gruppi.
Forme della rodopsina
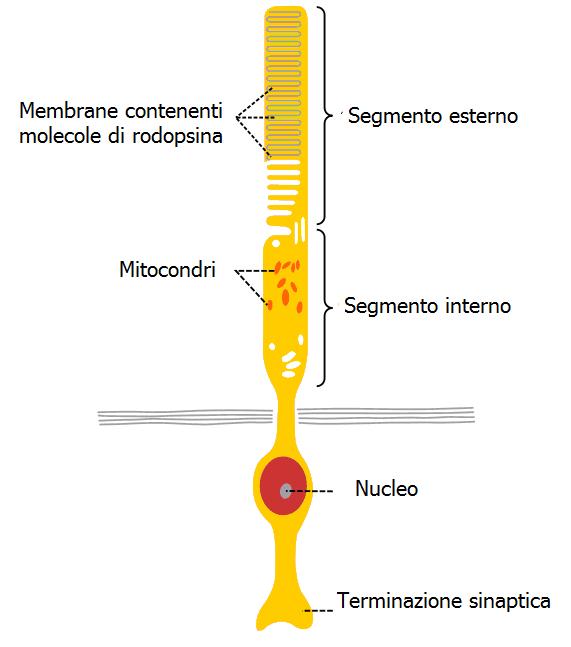
I fotorecettori consistono in un segmento esterno connesso mediante il ciglio al segmento interno contenente il nucleo della cellula. La porzione esterna racchiude diverse centinaia di sottili dischi membranosi (lamelle).
Nei bastoncelli le lamelle sono dischi separati dalla membrana cellulare, mentre nei coni consistono in una singola membrana ripiegata con quella plasmatica. La membrana delle lamelle contiene molecole di fotopigmento (rodopsina) un singolo bastoncello umano ne contiene 100 milioni. La rodopsina consiste in due parti: l’opsina (proteina) e il retinale (lipide) sintetizzato dalla vitamina A legato all’opsina mediante una base di Schiff.
Il retinale è una molecola a lunga catena che può esistere in due forme (isomeri); una forma a catena dritta (tutto-trans) e una forma ripiegata (11.cis). Il retinale 11 cis è l’unica forma che può legarsi all’opsina. Quando il retinale 11 cis assorbe un fotone di luce, la catena si raddrizza in forma tutto trans, un processo detto fotoisomerizzazione, e la molecola di fotopigmento si scinde nelle sue parti costitutive.(il pigmento è stato sbiancato).
Trasduzione visiva: proprietà elettriche e chimiche dei fotorecettori
In condizioni di oscurità, i bastoncelli e i coni hanno un potenziale di membrana a riposo di –40mV. Questo perché una continua corrente al buio fluisce nel segmento esterno quando gli ioni Na si spostano lungo il loro gradiente elettrochimico attraverso l’apertura dei canali cationici. La luce causa l’iperpolarizzazione della membrana cellulare, chiudendo i canali cationico nella membrana del segmento esterno. I canali cationici sono normalmente tenuti aperti dal guanosil 3-5 monofosfato ciclico (GMPc).
La fotoisomerizzazione della rodopsina innesca una reazione a catena che risulta in una rapida caduta dei livelli di GMPc., provocando la chiusura dei canali cationici.
Per tale ragione il GMPc agisce come messaggero interno alla cellula, trasferendo informazioni circa la rilevazione di luce dalle molecole di rodopsina.
Quando un fotone è assorbito da una molecola di rodopsina, il retinale cambia conformazione da 11 cis a tutto trans; la proteina a questo punto passa attraverso una serie di forme intermedie; come la metarodopsina che lega alla membrana del disco la proteina G detta transducina.
Allo stato inattivo la transducina è legata ad una molecola di guanosin-disfosfato (GDP). La metarodopsina catalizza lo scambio di una molecola di GDP legata alla trasducina con una di guanosin trifosfato (GTP) che dopo aver avuto altre trasformazioni si lega a un enzima detto fosfodiesterasi che può catalizzare l’idrolisi di GMPc . Tale sistema può consentire l’idrolisi di 400.000 molecole di GMPc entro 1 sec di assorbimento di un singolo fotone. La caduta dei livelli di GMPc causa la chiusura dei canali Na e l’iperpolarizzazione dei recettori.
In condizioni di oscurità, gli ioni calcio (Ca), come quelli per il sodio (Na), entrano nella cellula attraverso l’apertura dei canali cationici e sono espulsi per mezzo di una pompa elettrogenica calcio-sodio. Il processo di trasduzione visiva porta a una caduta della concentrazione intracellulare di GMPc e la seguente chiusura dei canali cationici blocca l’entrata degli ioni Ca, ma essi continuano ad essere pompati fuori. E’ stato dimostrato che i variabili livelli di Ca agiscono come un meccanismo di retroazione che accellera il recupero della cellula e consente l’adattamento alla luce. Per spiegare questa azione sono stati proposti tre meccanismi:
- Il Ca intracellulare altera l’azione della guanilito-ciclasi, l’enzima responsabile della sintesi di GMPc. Tale alterazione è mediata da una proteina che lega gli ioni Ca, denominata recoverina. Si suppone che tale proteina attivi la guanilato-ciclasi a bassi livelli di Ca. Perciò quando i livelli di Ca diminuiscono, l’attività della guanilato-ciclasi aumenta, provocando una maggiore concentrazione di GMPc permettendo la riapertura dei canali cationici.
- L’affinità dei canali cationici GMPc dipendenti sembra essere diminuita dal Ca. Pertanto quando i livelli di Ca diminuiscono, l’affinità dei canali per il GMPc aumenta compensando la caduta dei GMPc.
- In presenza di alti livelli di Ca la S-modulina allunga la vita della PDE attiva e inibisce la fosforilazione della rodopsina, pertanto tale meccanismo potrebbe verificarsi a livello dell’inattivazione dei pigmenti.
I fotorecettori a volte mandano dei segnali elettrici anche quando non c’è luce, producendo eventi elettrici non distinguibili da quelli evocati dalla luce. Questo rumore di fondo rende difficile vedere bene al buio. Però alcuni studi hanno scoperto che questo disturbo può anche aiutare a vedere dei segnali molto deboli grazie al meccanismo conosciuto come risonanza stocastica. La risonanza stocastica è un fenomeno in cui il rumore casuale amplifica le frequenze del segnale e lo rende più evidente.
Conclusione
In conclusione, grazie al processo di trasduzione visiva, le informazioni visive sono elaborate dai fotorecettori (coni e bastoncelli) che contengono i fotopigmenti (rodopsina e iodopsine). Questi fotopigmenti sono composti da una parte proteica (opsina) e da un cromoforo (retinale) che cambia conformazione quando assorbe un fotone, attivando una cascata intracellulare che porta alla chiusura dei canali cationici e all’iperpolarizzazione dei recettori.
Il calcio ha un ruolo di retroazione e di adattamento alla luce, modulando l’attività della fosfodiesterasi, della guanilato ciclasi e dei canali cationici. Successivamente, le cellule gangliari estraggono le informazioni dai fotorecettori.
Le cellule gangliari hanno una disposizione centro-periferia dei fotorecettori, che crea una risposta antagonista (ON/OFF) a seconda che la luce colpisca il centro o la periferia del loro campo recettivo. Questa risposta permette di rispondere bene a qualsiasi orientamento del bordo e di formare immagini nitide e dettagliate. Le cellule gangliari hanno una disposizione centro-periferia dei fotorecettori, che crea una risposta antagonista (ON/OFF) a seconda che la luce colpisca il centro o la periferia del loro campo recettivo. Questa risposta permette di rispondere bene a qualsiasi orientamento del bordo e di formare immagini nitide e dettagliate.
La retina comprime e riordina l’informazione dei fotorecettori per individuare le differenze di luce tra zone vicine, che indicano i bordi degli oggetti.