Il sistema nervoso utilizza due meccanismi fondamentali per la trasmissione sinaptica: la trasmissione elettrica e la trasmissione chimica.
Sinapsi elettrica e sinapsi chimica
Le sinapsi chimiche hanno caratteristiche morfologicamente diverse da quelle elettriche. A livello delle sinapsi chimiche non esiste continuità citoplasmatica fra le cellule, i neuroni sono separati da una fessura sinaptica. Mentre nelle sinapsi elettrice i esistono particolari canali comunicanti che stabiliscono un ponte tra il citoplasma delle due cellule. La corrente passando in questi canali incontra bassa resistenza ed elevata conduttanza, e quindi la corrente deposita cariche positive sulla membrana della cellula postsinaptica depolarizzandola.
Nelle sinapsi chimiche la corrente uscente nelle cellula presinaptica esce semplicemente all’esterno attraverso i canali passivi, e non tenderà ad attraversare la membrana della cellula postsinaptica che ha resistenza elevata. Il potenziale di azione che arriva nella terminazione di una sinapsi chimica determinerà invece la liberazione di una sostanza trasmettitrice che diffonderà attraverso la fessura sinaptica e andrà a legarsi con un recettore specifico che potrà depolarizzare o iperpolarizzare la cellula postsinaptica.
Le sinapsi elettriche
Le sinapsi elettriche si formano fra una fibra presinaptica di grandi dimensioni e un neurone postsinaptico piuttosto piccolo, in quanto per depolarizzare la cellula postisinaptica è necessaria una grande densità di corrente che deve provenire dai canali voltaggio dipendenti della cellula presinaptica. Questi canali infatti devono fornire una corrente sufficiente per determinare un’efficace modificazione del potenziale nella cellula postsinaptica. Per riuscire a far ciò le fibre presinaptiche devono essere piuttosto grandi.
La latenza tra il potenziale di azione presinaptico e il potenziale postsinaptico è particolarmente breve. Per innescare una risposta nella cellula postsinaptica non è necessaria la presenza di un potenziale di azione nella cellula presinaptica. Anche uno stimolo depolarizzante sotto soglia determina una depolarizzazione passiva nella cellula postsinaptica. Per questo motivo la sinapsi elettrica è chiamata spesso trasmissione elettrotonica. Inoltre tali sinapsi sono chiamate bidirezionali per via delle loro proprietà passive, esse infatti sono in grado di trasmettere con la stessa efficienza segnali dalla cellula presinaptica a quello post o in senso inverso.
Questo tipo di trasmissione riveste notevole importanza per tutte le risposte che mediano comportamenti di fuga e di pericolo. Aggregati interi di neuroni possono essere connessi con sinapsi elettriche. In questi casi la corrente che passa attraverso la membrana di una cellula potrà entrare o uscire attraverso la membrana di tutta la popolazione delle cellule elettricamente accoppiate. E’ in questo modo che parecchie cellule piccole possono esercitare funzioni che in generale spettano a cellule molto più grandi.

Nelle sinapsi elettrice, la zona di opposizione dei due neuroni è detta giunzione comunicante. Questo spazio è attraversato da speciali strutture proteiche, i canali delle giunzioni comunicanti, attraverso il quale passa il flusso delle correnti che provengono dal neurone presinaptico e vanno a quello postsinaptico. La conduttanza dei canali delle giunzioni comunicanti può variare ad es. per un abbassamento del pH o ad elevate concentrazioni di Ca. A livello di alcune giunzioni specializzate, i canali sono voltaggio sensibili, il che fa si che la trasmissione divenga unidirezionale. Infine, i neurotrasmettitori liberati dalle sinapsi chimiche poste in vicinanza possono attivare protein-chinasi che diffondono fino alle giunzioni comunicanti e modificano l’accesso dei canali.
Inoltre, dato il grande diametro del poro delle giunzioni, le sinapsi elettriche oltre a conferire velocità e sincronia del messaggio nervoso, possono trasmettere anche segnali metabolici. I pori infatti lasciano passare facilmente ogni tipo di catione ed anione ed anche composti organici come il secondo messaggero IP e l’AMP ciclico.
Tutti i canali delle giunzioni sono formati da due strutture a cilindro uno dei quali appartiene alla cellula presinaptica e l’altro a quella postsinaptica. I due cilindri vengono a mutuo contatto in modo da formare un canale che pone in comunicazione il citoplasma delle due cellule.
Le sinapsi chimiche

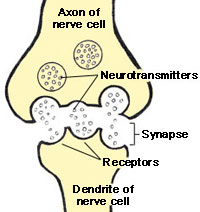
In queste sinapsi non esiste nessun elemento che connette le due cellule. Infatti la fessura sinaptica è un po’ più larga. Nelle terminazioni presinaptiche sono presenti agglomerazioni di particolari strutture vescicolari, le cosiddette vescicole sinaptiche, ciascuna delle quali contiene migliaia di neurotrasmettitori Queste sostanze vengono liberate nella fessura sinaptica in risposta all’ingresso di Ca. Una volta liberato, il neurotrasmettitore si diffonde nello spazio extracellulare e si lega a particolari siti localizzati sulla cellula postsinaptica, determinando l’apertura o la chiusura di particolari canali ionici.
Tutti questi passaggi giustificano la presenza di un ritardi sinaptico. Le sinapsi chimiche possiedono la caratteristica dell’amplificazione, infatti, la liberazione anche solo di una vescicola da luogo alla liberazione di migliaia di neurotrasmettitori che determinerà l’apertura di migliaia di canali ionici.
Questa liberazione diretta in maniera precisa viene effettuata a livello di strutture secretorie dette zone attive.
Lo stesso neurotrasmettitore è in grado di svolgere funzioni diverse; in alcuni gruppi di cellule viene liberato al livello delle zone attive e si comporta come un trasmettitore agendo sulle cellule bersaglio; in altre terminazioni esso può venire liberato in maniera meno precisa e fungere da modulatore determinando risposte più diffuse.
Tutta una serie di molecole a basso peso molecolare e di peptidi possono fungere da neurotrasmettitore, ma l’azione che essi esercitano sull’elemento postsinaptico dipende dalle proprietà del recettore.
Il concetto di recettore è stato introdotto alla fine dell’800 e si basa su due caratteristiche:
- Sono proteine che attraversano la membrana a tutto spessore. La zona che è in contatto con l’ambiente extracellulare ha la funzione di riconoscere e di legare il neurotrasmettitore
- Essi svolgono una funzione effettrice sulla cellula aprendo o chiudendo l’accesso dei canali
Le vie di accesso dei canali ionici possono essere dirette o indirette:
- I recettori che agiscono direttamente sull’accesso dei canali ionici sono costituiti da proteine integrali della membrana composte da diverse subunità polipeptidiche che formano un’unica macromolecola che comprende si l’elemento deputato al riconoscimento del neurotrasmettitore che il canale ionico. Questi recettori ionotropici quando si legano ad un neurotrasmettitore vanno incontro a una modificazione della propria conformazione.
- I recettori che regolano indirettamente l’accesso ai canali ionici (norepinefrina, serotonina), sono costituite da macromolecole distinte dai canali ionici sui quali agiscono. In questa categoria troviamo:
a) I recettori metabotropi. Si tratta di proteine integrali della membrana che si accoppiano con un gruppo di proteine che legano il GTP (proteine G). Queste proteine G attivano a loro volta un gruppo di enzimi effettori che catalizzano la formazione di una serie di secondi messaggeri come l’AMP ciclico. Questi secondi messaggeri possono agire direttamente sui canali ionici o attivare una particolare famiglia di enzimi come la protein ciclasi.
b) I recettori che appartengono alla famiglia delle tirosin-chinasi. Queste chinasi fosforilano residui di tirosina modulando l’accesso ai canali.
Queste canali indiretti sono molto lenti e servono a modulare le manifestazioni del comportamento.
Nel sistema nervoso la forma preminente di trasmissione sinaptica è quella chimica che ha il vantaggio di amplificare i segnali ed è modulabile.
