La corteccia visiva primaria (V1), o corteccia striata, è un’importante area in cui le informazioni in uscita dalla retina e dal LGN (nucleo genicolato laterale), parzialmente elaborate, sono separate e poi categorizzate per un’analisi più approfondita nelle aree visive specializzate della corteccia extrastriata. La maggior parte dei neuroni ha proprietà di risposta che cambiano notevolmente man mano che si procede nello studio di V1; dalla sensibilità a barre o linee di diverso orientamento o in movimento in diverse direzioni alla specializzazione nel colore. Inoltre, l’organizzazione in moduli e colonne che si ripetono sembra essere una configurazione comune a tutte le aree visive corticali ed è un sistema efficiente per rappresentare uno stimolo multidimensionale su una regione della corteccia bidimensionale.
Organizzazione della corteccia visiva primaria (V1)
La corteccia visiva primaria (V1) è situata nella parte posteriore del lobo occipitale ed è raggiunta dalle informazioni visive attraverso le radiazioni ottiche che giungono dal LGN. V1 può essere riconosciuta dal suo particolare aspetto: fasci di fibre afferenti formano, in quest’area, delle strisce chiare, da cui deriva il nome di corteccia striata. L’area che circonda immediatamente V1 è V2 (area 18) e riceve segnali in entrata principalmente da V1.
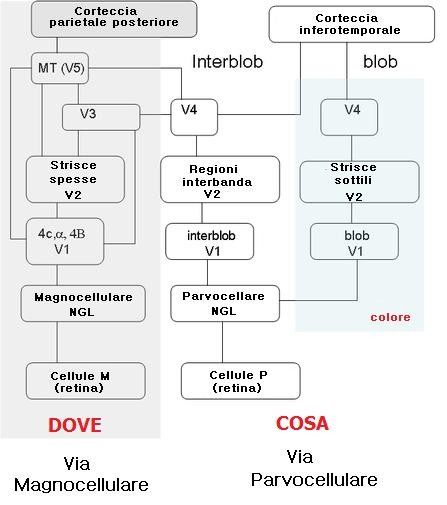
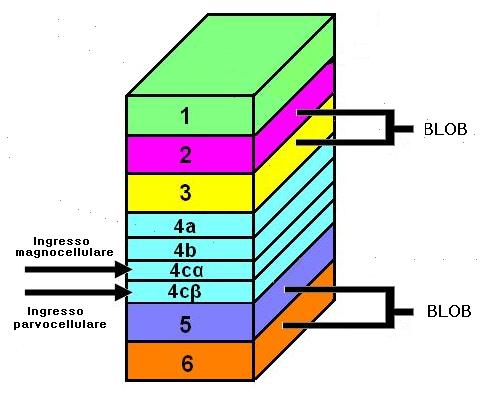
La topografia del campo visivo è conservata dalle proiezioni dall’LGN a V1: qui la fovea è rappresentata in modo maggiore di quella dedicata al campo visivo periferico. In V1 La maggior parte delle fibre proveniente dal LGN terminano nello strato 4 che è suddiviso in tre sottostrati: A, B, C. Il C è ulteriormente suddiviso in 4Cα e 4Cβ.
Le proiezioni degli strati parvocellulari (P) del LGN terminano nei substrati 4A, 4Cβ e nella parte superiore dello strato 6. Le cellule nello strato 4Cβ proiettano ai neuroni degli strati 2, e 3.
Gli strati magnocellulari (M) terminano in 4Cα e nella parte inferiore dello strato 6. Le cellule di 4Cα proiettano allo strato 4B. Entrambe le M e P mandano segnali ai blobs.
Per maggiori informazioni sull’organizzazione del sistema visivo.
Colonne di dominanza oculare
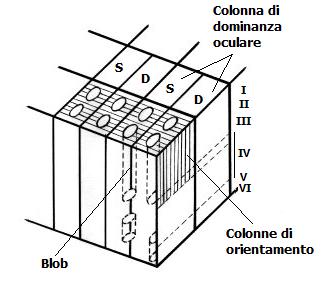
I segnali provenenti dai due occhi sono mantenuti separati nello strato 4. Infatti, le cellule di uno strato del LGN proiettano a un gruppo di cellule bersaglio nello strato 4C, separato dai gruppi a cui giungono i segnali dall’altro occhio. Al di sotto e al di sopra di questo strato, la maggior parte delle cellule riceve informazioni da entrambi gli occhi, sebbene un occhio sia generalmente dominante. Hubel e Wiesel hanno nominato queste colonne di cellule colonne di dominanza oculare. I blobs, selettivi per il colore sono localizzati nel centro di ogni colonna di dominanza oculare.
Le cellule centro-on e centro off, le cellule semplici e cellule complesse
Le connesioni che si stabiliscono all’interno di V1 trasformano le informazioni in modo tale che la maggior parte delle cellule degli strati soprastanti lo strato IV rispondono in modo più selettivo a stimoli nettamente più complessi.
Hubel e Wiesel osservarono che le cellule di tutti gli strati eccetuate quelle dei blob rispondono in maniera ottimale a stimoli di forma lineare come un segmento o una barretta.
Gli autori hanno suddiviso queste cellule in due categorie, cellule semplici e cellule complesse.
Esiste inoltre, una classe di cellule osservata esclusivamente nello strato 4C avente un campo recettivo con organizzazione centro-periferia, come quello delle cellule dell’LGN e delle gangliari retiniche.
Le cellule semplici
Le cellule semplici si trovano prevalentemente negli strati 4 e 6, e ricevano segnali in entrata direttamente dall’LGN. Un tipo di cellule semplici possiede un campo recettivo che consiste in una stretta regione centrale fiancheggiata da due aree antagoniste. Il centro può essere eccitatorio o inibitorio. Per questo tipo di cellule, l’attivazione ottimale è costituita da una barra di luce di dimensioni pari a quelle dell’area centrale e che abbia un determinato orientamento Essa riceve afferenze da diverse cellule dell’LGN con organizzazione antagonistica centro-periferia che appunto determina l’ampiezza e l’orientamento ottimale della barra luminosa o scura. La risoluzione non è perduta ma incorporata in un campo recettivo più complesso.
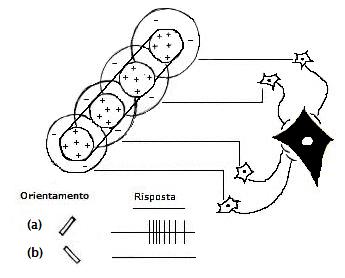
In V1 le cellule hanno campi recettivi rettilinei con distinte zone eccitatorie ed inibitorie. Per semplicità, sono rappresentati campi recettivi con asse di orientamento verticale. Alcuni neuroni hanno una zona centrale eccitatoria circondata da una inibitoria (1), altre hanno una disposizione opposta (2,3) o presentano una zona eccitatoria a fianco di una inibitoria (4).
Perciò lo stimolo avrà la massima efficacia quando il suo orientamento sarà perfettamente coincidente con quello del campo recettivo della cellula stessa. Lo stimolo, perciò deve occupare soltanto la zona eccitatoria del campo recettivo e non deve sconfinare nella regione inibitoria.
Le diverse cellule corticali che ricevono le proprie afferenze dallo stesso punto della retina hanno campi recettivi simili ma caratterizzati da assi di orientamento diversi. In tal modo per ogni punto della retina saranno presentati in corteccia tutti gli assi di orientamento.
Si ritiene che i campi recettivi lineari celle cellule semplici prendono origine dalla loro disposizione, in quanto gruppi di cellule stellate aventi analoghe proprietà di risposta si dispongono ordinatamente nei sottostrati IVC.
Le cellule complesse
Anche le cellule complesse sono cellule piramidali ma sono localizzate in zone più lontane dai sottostrati d’ingresso. La maggior parte di queste cellule è localizzata negli strati II, III, V e VI. Anche loro possiedono campi recettivi lineari con assi di orietamento specifici. I loro campi recettivi però hanno dimensioni maggiori e non possiedono zone ecciatatorie ed inibitorie ben definite e la posizione esatta dello stimolo non appare essenziale. Per alcune di queste cellule anche il movimento sembra rappresentare uno stimolo efficace.
Le cellule semplici e quelle complesse possono rivestire un ruolo importante per l’analisi dei contorni di una immagine, come la sua forma, ma anche ai suoi contrasti di luminosità ma non all’interno degli oggetti. Le parti interne degli oggetti sono monotone e non contengono alcuna informazione visiva. E’ invece l’informazione che arriva ai margini che ci permette di riconoscere i diversi oggetti.
Alcune cellule complesse e semplici possiedono dei margini di arresto. La frequenza di scarica di queste cellule aumenta all’’aumentare della lunghezza dello stimolo luminoso all’interno del campo recettivo soltanto entro specifici limiti; l’allungamento della baretta al di là di una certa estensione in entrambe le direzioni riduce ed, infine, annulla la risposta cellulare. Questa limitazione può verificarsi in una sola direzione (cellule ad margine d’arresto singole) o in entrambi le direzioni (cellule a doppio margine d’arresto).
Queste cellule possono essere immaginate come formate dalla convergenza di tre cellule aventi proprietà comuni. In cui due inibitorie sono poste a fiancheggiare la cellula che ha campo recittovo eccitatorio.
Per una cellula a margine d’arresto singolo lo stimolo ottimale è costituito da una barretta lineare che s’interrompe bruscamente.
Per una cellula a doppio margine di arresto gli stimoli più efficaci sono rappresentati da brevi segmenti lineari.
La funzione di queste cellule consiste nel valutare le lunghezze di un tratto lineare, e perciò sono in grado di riconoscere angoli, le curvature o le brusche interruzioni di un margine.
Selettività di risposta dei neuroni in V1

La frequenza spaziale
Sebbene molti neuroni in V1 rispondano a linee o barre, lo stimolo ottimale è un reticolo sinusoidale che appare come una serie di barre parallele indistinte e sfuocate. Un’onda sinusoidale è classificata in base alla sua frequenza spaziale. Dato che le dimensioni dell’immagine di uno stimolo dipende dalla sua distanza rispetto all’occhio, l’unità di misura utilizzata è l’angolo visivo, al posto della distanza fisica tra i cicli adiacenti. La maggior parte dei neuroni della corteccia striata risponde in modo preferenziale a un reticolo sinusoidale di particolare frequenza spaziale, posto in una parte appropriata del campo visivo.
La tessitura
Recentemente è stata identificata in V1, una nuova classe di neuroni che sembra essere selettivo per la tessitura che rispondono a reticoli sinusoidali o a reticoli d’onda quadra aventi particolare frequenza spaziale e orientamento. Esse non sembrano analizzare la frequenza spaziale come le precedenti ma la tessitura dell’immagine e potrebbero contribuire alla percezione della profondità.
La selettività per la direzione del movimento
Circa il 10% – 20% delle cellule complesse, negli strati superiori della corteccia striata, mostrano una forte selettività per la direzione dello stimolo. Questo tipo di cellule risponde fortemente al movimento in una particolare direzione, mentre è silente al movimento in altre direzioni.
Barlow e Levick hanno proposto un modello di attivazione per le cellule selettive alla direzione del movimento nella retina del coniglio. Gli autori hanno supposto che tra le cellule semplici e complesse ci siano cellule di tipo intermedio. Ogni cellula intermedia è eccitata da una cellula semplice e inibita da una cellula semplice per mezzo di un’altra cellula intermedia. Se uno stimolo si muove in una certa direzione (direzione nulla), la prima cellula intermedia è eccitata dalla cellula semplice, ma inibita dalla seconda cellula intermedia cancellando quindi l’eccitazione, se invece lo stimolo si muove in direzione opposta, il segnale inibitorio arriva troppo tardi.
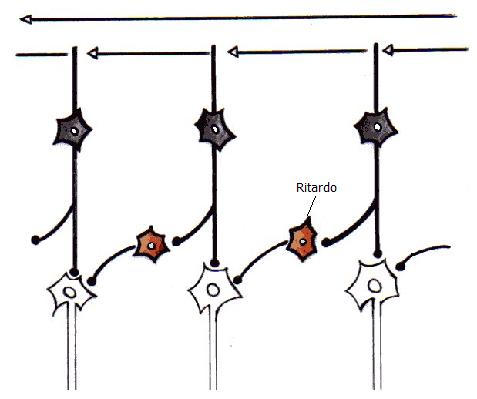
Le tre cellule bianche convergono su una cellula superiore.
Un esempio di adattamento selettivo è dato dal fenomeno della post-immagine da movimento (illusione a cascata, per un esempio vedi qui). Se si osserva uno stimolo in movimento verso una data direzione per un certo tempo, come un reticolo o una cascata, e poi si fissa un oggetto statico, questo sembrerà muoversi in direzione opposta a quello stimolo. Questo perché una prolungata esposizione a un movimento verso il basso affaticherà o adatterà le cellule sensibili a tale movimento, e conseguentemente non manifesteranno alcuna attività spontanea. Le cellule sensibili al movimento verso l’alto invece, avranno normali livelli di attività spontanea, e questo squilibrio produrrà una configurazione di attività simile a quella prodotta dal movimento verso l’alto.
Per maggiori informazioni sulla percezione del movimento.
Il colore
Ogni blob è centrato all’interno di ogni colonna di dominanza oculare. All’interno di ogni colonna il grado di dominanza varia. Nel centro di ogni colonna, la dominanza sarà assoluta pertanto i neuroni riceveranno segnali solo da un occhio. I blob coincidono con questi centri di monocularità. Essi contengono cellule ad opponenza semplice e doppia.
All’interno di ogni blob i neuroni avranno un’opponenza rosso/verde o giallo/blu, le due forme di opponenza non sono mescolate nello stesso blob. Queste osservazioni suggeriscono che ogni singolo blob è dedicato all’analisi di un solo sistema di opponenza cromatica. Inoltre, i diversi tipi di blob non sono equamente distribuiti in V1.
La retina contiene una maggior quantità di coni verdi e rossi rispetto ai blu.
I blob giallo/blu sembrano, essere raggruppati insieme suggerendo che V1 non riceve segnali cromatici giallo/blu in modo uniforme. Questo sarebbe in accordo con l’organizzazione anulare dei coni blu nella retina. I blob spesso sembrano essere appaiati. Il blob in una particolare colonna di dominanza oculare è connesso, mediante ponti al blob sito nella colonna oculare vicina. I ponti connettono blobs di diversa opponenza cromatica e le loro cellule non rispondono né al rosso/verde né al giallo/blu ma presentano una selettività spettrale combinata.
Per maggiori informazioni sulla percezione del colore.
L’organizzazione combinata
Diverse aree visive sembrano essere suddivise in moduli, o super colonne, di analisi che ricevono informazioni da altri moduli, Si suppone che V1 comprenda 2500 moduli. I neuroni in ogni modulo sono coinvolti nell’analisi di una particolare porzione del campo visivo.
Nel modello del cubo di ghiaccio; la corteccia visiva primaria è divisa in moduli contenenti due blob e regioni interblob. La corteccia è divisa in colonne di dominanza oculare e perpendicolarmente a queste si dispongono le colonne di orientamento. I neuroni degli strati 4Cα e 4Cβ sono monoculari. Ogni unità (CDO) riceve segnali da un occhio solo. Le due unità però si scambiano informazioni, mentre l’80% dei neuroni negli altri strati sono binoculari. Perciò ogni modulo (super colonna) contiene neuroni sensibili alle lunghezze d’onda, al movimento, alle linee o bordi, di particolare orientamento entro una specifica regione del campo visivo. Man mano che si sposta parallelamente alla superficie corticale, l’orientamento per ogni neurone varia progressivamente di 10 gradi. Muovendosi attraverso le due metà del modulo, è possibile osservare insiemi di rotazioni di 180 gradi.
Con penetrazioni verticale di un microelettrodo, l’orientamento è chiaramente definito e costante negli strati sopra e sotto 4C( prive di orientamento). Nella penetrazione obliqua, si osserva una progressiva e sistematica variazione di 10 gradi nella preferenza di orientamento.
Attraverso la tecnica dell’optical imaging è stato dimostrato però che ci sono anche discontinuità e distorsioni. Ciò potrebbe essere dovuto dal fatto che sono necessarie discontinuità per preservare le relazioni retinotipiche sulla corteccia corticale bidimensionale.
Mi piacerebbe conoscere il tipo di stimolo dei neuroni sensibili al movimento. E’ esso uno stimolo spazio-temporale o più semplicemente del tipo frequenza doppler?
Hubel e Wiesel nei loro studi hanno utilizzato stimoli luminosi proiettati all’interno del campo visivo dell’animale. Per un esempio può vedere questo video dove si possono osservare le rispooste delle cellule semplici e di quelle complesse
l’effetto è chiaro , sarebbe interessante conoscere la causa.Grazie!
Questo è un articolo ?
No, non è un loro articolo. Si tratta di una spiegazione dell’architettura e delle funzioni della corteccia visiva che ho scritto riprendendo le spiegazioni e i libri universitari.
Scusatela domanda, Questo è un articolo ufficiale di hubel e Wiesel?