I sistemi di secondo messaggero hanno alla base molte caratteristiche in comune. Il legame di un neurotrasmettitore al recettore attiva una proteina G trasduttrice capace di attivare enzimi che fungono da effettori primari. Questi enzimi danno origine alla formazione di un secondo messaggero, che può attivare un effettore secondario o agire direttamente su proteine bersaglio regolatrici.
I recettori ionotropici comprendono due famiglie di geni:
- la prima famiglia comprende i recettori per l’acetilcolina (Ach), per l’acido aminobutirrico (GABA), per la glicina e la serotonina.
- La seconda invece comprende due classi di recettori per il glutammato: l’NMDA e i recettori non-NMDA.
I recettori metabotropici che controllano indirettamente i canali si possono distinguere in:
- recettori metabotropici in cui la molecola recettrice è accoppiata alla molecole effettrice da una proteina (proteina G) che si lega ad un nucleotide della guanosina, Per questo motivo questi recettori sono noti anche come recettori accoppiati a proteine G. Questa famiglia comprende i recettori adrenergici, i recettori muscarinici (Ach), una classe di recettori per il GABA, il glutammato e la serotonina, i recettori dei neuropeptidi e la rodopsina. In generale l’effettore è costituito da un enzima che produce un secondo-messaggero diffusibile. I secondi-messaggeri a loro volta innescano a cascata una serie di reazioni biochimiche, che vanno ad attivare protein-chinasi specifiche in grado di fosforilare diverse proteine cellulari.
- Anche la seconda famiglia di recettori che agiscono indirettamente sui canali ionici comprende numerosi membri, dei quali il più conosciuto è la tirosin-chinasi. Questo è un enzima capace di autofosforilarsi o di fosforilare altre proteine.
- Un’altra classe di secondi-messaggeri è prodotta dall’idrolisi dei fosfolipidi delle membrane cellulari e quella costituita da sostanze gassose facilmente diffusibili, per es. NO e il CO.
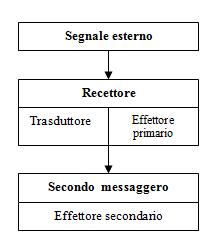
Le singole vie metaboliche di secondo messaggero hanno alla base molte caratteristiche in comune. A differenza dei recettori che regolano direttamente l’accesso dei canali, i recettori accoppiati alle proteine G sono formati da un’unica subunità che comprende 7 segmenti. Il legame di un neurotrasmettitore a un recettore costituito da 7 segmenti di catena polipeptidica attiva una proteina G trasduttrice capace di attivare enzimi che fungono da effettori primari. Questi enzimi danno origine alla formazione di un secondo messaggero, che può attivare un effettore secondario o agire direttamente su proteine bersaglio (regolatrici).
Via dell’AMP-ciclico
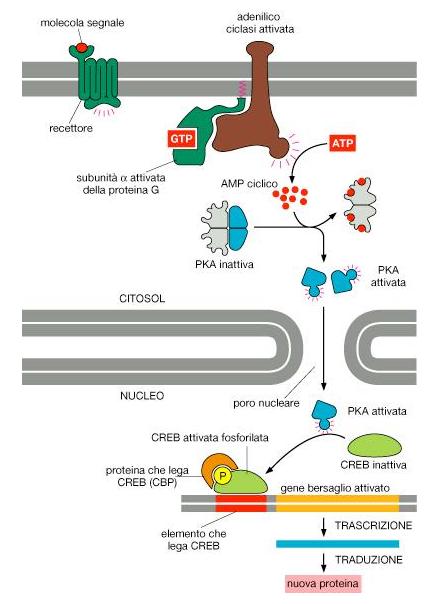
Il legame di un neurotrasmettitore al recettore determina l’attivazione di una proteina G trasduttrice. Quando è a riposo, allo stato inattivo G è normalmente legata ad una molecola di GDP.
Quando viene attivata G prende una molecola di GTP in scambio del GDP ed attiva l’enzima adenill-ciclasi (effettore primario).
Questo enzima è una proteina integrale della membrana che catalizza la conversione di ATP in AMPc (secondo messaggero).
L’idrolisi del GTP ne delimita il distacco dalla ciclasi che diviene inattivata. Il recettore e la ciclasi perciò non agiscono direttamente ma sono accoppiati mediante la proteina G.
L’AMPc attiva le protein chinasi AMPc-dipendente (effettore secondario) determinando la dissociazione delle sue subunità regolatrici dalle sue subunità catalitiche. In questo modo queste ultime saranno in grado di fosforilare specifici substrati proteici.
Quando la concentrazione di AMPc tende a diminuire all’interno della cellula le subunità regolatrici libere si associano nuovamente con le subunità catalitiche. L’aumento della concentrazione di AMP ciclico attiva la protein chinasi A (PKA) nel citosol, e le subunita’ catalitiche che vengono rilasciate migrano nel nucleo, dove fosforilano la proteina regolatrice CREB.
Una volta fosforilata, CREB richiama il coattivatore CBP, che stimola la trascrizione genica.
Un meccanismo simile si ha quando i secondi messaggeri sono formati dall’idrolisi dei fosfolipidi della membrana determinata dall’attivazione di due enzimi: la fosfolipasi A e la fosfolipasi C .
Sistema del diacilglicerolo – l’inositol-tridosfato.
L’attivazione della fosfolipasi C avviene da parte una proteina G che è stata a sua volta attivata dal legame del neurotrasmettitore col recettore. Questa fosfolipasi degrada il fosfatidil-inositolo (PIP2) in due secondi messaggeri : l’inositol-tridosfato (IP3) e al diaciglicerolo (DAG)
Protein-chinasi C.
Il diacilglicerolo attiva la protein-chinasi C (PKC) che è in grado di fosforilare molti substrati proteici. Un aspetto importante che deriva dalla fosfolipasi C è costituita dal fatto che l’ IP3 può far aumentare la concentrazione di Ca all’interno della cellula. I Ca agiscono spesso solo dopo aver formato un complesso con una molecola di Calmodulina. Un esempio può essere l’attivazione della protein chinasi Ca/calmodulina-dipendente.
Via dell’acido arachidonico.
L’acido arachidonico è liberato in seguito all’attivazione della fosfolipasi A (PLA) che idrolizza il fosfoinositolo (PI). L’acido arachidonico può essere metabolizzato attraverso tre enzimi: dalle clossigenasi (che producono le prostaglandine e possono essere inibiti dai farmaci anti-infiammatori non stereoidei come l’aspirina), dalle lipossigenasi e dal complesso contenente la famiglia del citocromo P450.
Funzioni dei sistemi di secondo messaggero
In generale, i sistemi di secondo messaggero alterano l’attività dei canali ionici fosforilando la proteina che ne costituisce il canale. La fosforilazione promossa dalle chinasi dei secondi messaggeri esercita alcuni effetti:
- Essa può innanzitutto aprire canali che sono normalmente chiusi al potenziale di membrana di riposo. In alcune cellule alcuni canali potassio (K) passivi che contribuiscono a determinare il livello di potenziale di riposo sono sotto il controllo di secondi messaggeri. La chiusura di questi canali esercita un’azione depolarizzante sui neuroni Un esempio può essere costituito dai recettori muscarinici presenti nei neuroni dei gangli simpatici e in certi neuroni dell’ippocampo e della corteccia cerebrale. Essi attivano un sistema di secondo messaggero che chiude un canale K voltaggio dipendente detto canale M. La chiusura di alcuni canali K farà diminuire l’efflusso continuo di K e determinerà un accumulo di cariche positive nella cellula. L’attività dei secondi messaggeri non si esercita soltanto sui canali passivi. Alcuni canali voltaggio dipendenti vengono chiusi da secondi messaggeri.
- Oltre a modulare la chiusura di canali che sono aperti al potenziale di riposo, i secondi messaggeri, possono diffondere all’interno del citoplasma e possono quindi esercitare i propri effetti su parti lontane dai recettori.
- L’andamento temporale delle attività sinaptiche mediate da secondi messaggeri è molto più lento, quindi ad es. possono produrre un potenziale eccitatorio lento di lunghissima durata..
Come si deduce da questi esempi le attività sinaptiche prodotte dai secondi messaggeri, in generale modulano l’eccitabilità dei singoli neuroni, mentre le attività sinaptiche dirette mediano la trasmissione sinaptica da un neurone ad un altro.
In alcune cellule uno stesso canale può essere modulato in due modi diversi in risposta all’impatto con un neurotrasmettitore: direttamente in maniera rapida, legandosi con una proteina G, e indirettamente con andamento temporale più lento, venendo fosforilato.
Un secondo messaggero inoltre può regolare l’efficacia dei recettori che attivano il proprio sistema di secondo messaggero e la sensibilità del recettore rendendolo ad esempio refrattario all’applicazione del proprio neurotrasmettitore. Questo processo viene detto desensitizzazione e dipende dai processi di fosforilazione.
Infine, come visto nel caso della via AMP-ciclico esiste un tipo di attività sinaptica attraverso il quale i secondi messaggeri fosforilano i processi di trascrizione delle proteine che regolano i processi dell’espressione genetica. Questo tipo di attività può innescare modificazioni che possono durare giorni o anche più.